第九章索引
第一节 概述
一、感受器、感觉器官的定义和分类
感受器是指分布在体表或组织内部的一些专门感受机体内、外环境改变的结构或装置。感受器的组成形成是多种多样的:有些感受器就是外周感觉神经末稍本身,如体表或组织内部与痛觉感受有关的游离神经末稍;有的感受器是裸露在神经末稍周围再包绕一些特殊的、由结缔组织构成的被膜样结构;但是对于一些与机体生存密切相关的感觉来说,体内存在着一些结构和功能上都高度分化了的感受细胞,它们以类似突触的形式直接或单位同感觉神经末稍相联系,如视网膜中的视杆和视锥细胞是光感受细胞,耳蜗中的毛细胞是声波感受细胞等,这些感受细胞连同它们的非神经性附属结构,构成了各种复杂的感觉器官如眼、耳等。高等动物中最重要的感觉器官,如眼、耳、前庭、嗅、味等器官,都分布在头部,称为特殊感管。
机体众多的感受器有不同的方法来分类。如根据感受器的分布部位,可分为内感受器和外感受器;根据感受器所接受刺激的性质,可分为光感受器、机械感受器、温度感受器和化学感受器等;便更常用的是结合刺激物和它们所引起的感觉或效应的性质来分类,据此所能区分出的人体的主要感觉类型和相应的感受器如表9-1所示。
表9-1 人体的主要感觉类型
| 感觉类型 |
感受器结构 |
感觉类型 |
感受器结构 |
| 视觉 |
视杆和视锥细胞 |
关节位置和运动觉 |
神经末稍 |
| 听觉 |
毛细胞 |
肌肉长度 |
神经末稍(肌梭) |
| 嗅觉 |
嗅神经元 |
肌肉张力 |
神经末稍(腱器官) |
| 味觉 |
味感受细胞 |
动脉血压 |
神经末稍 |
| 旋转加速度 |
毛细胞(三半规管) |
肺扩张 |
神经末稍 |
| 直线加速度 |
毛细胞(椭圆囊和球囊) |
头部血液温度 |
下丘脑某些神经元 |
| 触-压觉 |
神经末稍 |
动脉氧分压 |
神经末稍(?) |
| 温觉 |
神经末稍 |
脑脊液pH值 |
延髓腹外侧区感受器 |
| 冷觉 |
神经末稍 |
血浆葡萄糖 |
下丘脑某些细胞 |
| 痛觉 |
游离神经末稍 |
血浆渗透压 |
下丘脑前部某些细胞 |
表中只有前11项是通常能引起主观感觉的,其余的感受器一般只是向中枢神经系统提供内、外环境中某些因素改变的信息,引起各种调节性反应,但这时在主观上并不产生特定的感觉。
二、感受器的一般生理特性
(一)感受器官适宜刺激
各种感受器的一个共同功能特点,是它们各有自己最敏感、最容易接受的刺激形式;这就是说,用某种能量形式的刺激作用于某种感受器时,只需要极小的强度(即感觉阈值)就能引起相应的感觉。这一刺激形式或种类,就称为该感受器的适宜刺激,如在一定波长的电磁波是视网膜光感受细胞的适宜刺激,一定频率的机械震动是蜗毛细胞的适应刺激等。正因为如此,机体内、外环境中所发生的各种形式的变化,总是先作用于和它们相对应的那种感受器。这一现象的存在,是因为动物在长期的进化过程中逐步形成了具有各种特殊结构和功能的感受器以及相应的附属结构的结果,便得它们有可能对内、外环境中某些有意义的变化进入灵敏的感受和精确的分析。不同动物所处的生活环境和条件不同,因此在进化中有可能形成一些异于人体的特殊感受装置,这在广大的动物界屡见不鲜,早已引起人们极大的兴趣和注意。研究这些可能是极低等动物的特殊感受装置,不仅对理解感受器活动的一般规律有帮助,而且有很大的仿生学意义。
(二)感受器的换能作用
各种感受器在功能上的另一个共同特点,是能把作用于它们的各种刺激形式,转变成为相应的传入神经末稍或感受细胞的电反应,前者称为发生器电位(generator potential),在后者称为感受器电位(receptorpotential)。如在第二章所述,发生器电位和感受器电位的出现,实际上是传入纤维的膜或感受细胞的膜进行了跨膜信号传递或转换过程的结果。和体内一般细胞一样,所有感受器细胞对外来不同刺激信号的跨膜转换,也主要是通过两种基本方式进行的,如声波振动的感受与蜗毛顶部膜中与听毛受力有关的机械细胞对外来中与听毛受力有关的机械门控通道的开放和关闭有关,这使毛细胞出现与声波振动相一致的感受器电位(即微音器电位);视杆和视锥细胞则是由于它们的外段结构中视盘膜上存在有受体蛋白(如视紫红质),它们在吸收光子后,再通过特殊的G-蛋白和作为效应器酶的磷酸二酯酶的作用,引起光感受器细胞外段胞浆中cGMP的分解,最后使外段膜出现感受器电位。在其他一些研究过的感受器,也看到了类似的两种信号转换机制。由此可见,所有感受性神经末稍和感受器细胞出现电位变化,就是通过跨膜信号转换,把不同能量形式的外界刺激都转换成跨膜电位化的结果。
如前,发生器电位和感受器电位同终板电位和突触后电位一样,是一种过渡性慢电位,它们不具有“全或无”的特性而其幅度与外界刺激强度成比例;它不能作远距离传播而可能在局部实现时间性总和和空间性总和。正因为如此,感受器电位和发生器电位的幅度、持续时间和波动方向,就反映了外界刺激的某些特征,也就是说,外界刺激信号所携带的信息,也在换能过程中转移到了这种过渡性电变化的可变动的参数之中。
发生器电位和感受器电位的产生并不意味着感受器功能作用的完成,只有当这些过渡性电变化最终触发分布在该感受器的传入神经纤维上产生“全或无”式的可作远距离传导的动作电位序列时,才标志着这一感受器或感觉器官作用的完成;但主观感受的产生、对外界刺激信号的精细分析以及最后引起整个机体出现应答性反应和信息贮存等过程,则是传入神经纤维所输的神经信号到达各级脑中枢以后的反应,不属本章的内容。
(三)感受器的编码作用
感受器在把外界刺激转换成神经动作电位时,不仅仅是发生了能量形式的转换;更重要的是把刺激所包涵的环境变化的信息,也转移到了新的电信号系统即动作电位的序列之中,即编码作用。编码(encoding)一词,本是工程通讯理论中的一个概念,指一种信号系统(如莫尔斯电码)如何把一定的信息内容(如电文内容)包涵在少量特定信号的排列组合之中。因此,感受器将外界刺激转变成神经动作电位的序列时,同时也实现了编码作用;中枢就是根据这些电信号序列才获得对外在世界的认识的。问题是外界刺激的质和量以及其他属性,是如何编码在特有的电信号序列中的?这一问题十分复杂,目前还远远没有弄清楚,下面先从最简单的方面加以叙述。
首先考虑外界刺激的“质”,如听觉或视觉等刺激在性质上的不同是如何编码的。如所周知,不论来自何种感受器的传入神经纤维上的传入冲动,都是一些在波形和产生原理上基本相同的动作电位;例如,由视神经、听神经或皮肤感觉神经的单一纤维上记录到的动作电位,并无本质上的差别。因此,不同性质的外界刺激不可能是通过某些特异的动作电位波形或强度特性来编码的。实验和临床经验都表明,不同种类的感觉的引起,不但决定于刺激的性质和被刺激的感受器,也决定于传入冲动所到达的大脑皮层的终端部位。例如,用电刺激作用于病人视神经,使它人为地产生传向枕叶皮层的传入冲动,或者直接刺激枕叶皮层使之产生兴奋,这时都会引起光亮的感觉,而且主观上感到这些感觉是发生在视野的某一部位;同样,临床上遇到肿瘤或炎症等病变刺激听神经时,会产生耳鸣的症状,这是由于病变刺激引起的神经冲动传到了皮层听觉中枢所致;而某些痛觉传导路或相应中枢的刺激性病变,也会引起身体一定部位的疼痛。这些都说明,感觉的性质决定于传入冲动所到达的高级中枢的部位,而不是由于动作电位的波形或序列特性有什么不同;也就是说,不同性质的感觉的引起,首先是由传输某些电信号所使用的通路来决定的,即由某一专用路线(labeled line)传到特定终端部位的电信号,通常就引起某种性质的主观感觉。事实上,即使是同一性质的刺激范围内,它们的一些次级属性(如视觉刺激中不同波长的光线和听觉刺激中不同频率的振动等)也都有特殊分化了的感受器和专用传入途径。在自然状态下,由于感受器细胞在进化过程中的高度分化,使得某一感受细胞变得对某种性质的刺激或其属性十分敏感,而由此产生的传入信号又只能循特定的途径到达特定的皮层结构,引起特定性质的感觉。因此,一般无需怀疑,某种主观感觉是否是由一些非适宜刺激引起的“非真实”的感觉,只是在病理情况下有例外。
在同一感受系统或感觉类型的范围内,外界刺激的量或强度是怎样编码的呢?既然动作电位是“全或无”式的,因而刺激的强度不可能通过动作电位的幅度大小或波形改变来编码。根据在多数感受器实验中得到的实验资料,刺激的强度是通过单一神经纤维上冲动的频率高低和参加这一信息传输的神经张纤维的数目的多少来编码的。图9-1表示在人手皮肤的触压感受器所进行的实验,说明在感受器的触压重量和相应的传入纤维的动作电位发放频率之间,存在着某种对应关系。重量过轻时,神经纤维全无反应,到达感受阈值时开始有冲动产生;以后随着触压重量的增大,传入纤维上的冲动频率也越来越高。不仅如此,在触压刺激继续加大的情况下,同一刺激有可能引起较大面积的皮肤变形,使一个以上的感受器和传入纤维向中枢发放冲动。这样,刺激的强度既可通过每一条传入纤维上冲动频率的高低来反映,还可通过参与电信号传输的神经纤维的数目的多少来反映。当然,任何一个天然刺激在空间和时间上的属性都是极其复杂的(例如一个彩色电影画面所包涵的信息内容),因此,感受器的编码过程也是极其复杂的。还应该知道的是,感觉过程的编码过程并不只是感受器部位进行一次,事实上信息每通过一次神经元间的突触传递,都要进行一次重新编码,这使它有可能接受来自其他信息源的影响,使信息得到不断的处理,这当然属于中枢神经元网络的功能。
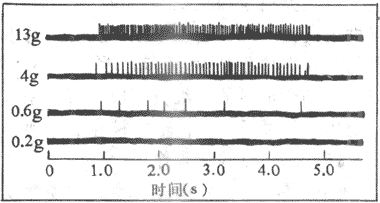
图9-1 不同重量的触压刺激是在单一传入纤维上引起的冲动频率的改变
至于刺激的物理强度如何转变成为传入神经纤维上频率不同的冲动,目前认为是由于强的刺激能引起幅度较大而持续时间较长的发生器电位,而后者引起神经末稍较高频率的冲动。
(四)感受器的适应现象
当刺激作用于感觉器时,经常看到的情况是虽然刺激仍在继续作用,但传入神经纤维的冲动频率已开始下降,这一现象称为感受器的适应(adaptation)。适应是所有感受器的一个功能特点,但它出现的快慢在不同感受器有很大的差别,通常可把它们区分为快适应和慢适应感受器两类。快适应感受器以皮肤触觉感受器为代表,当他们受刺激时只顾刺激开始后的短时间内有传入冲动发放,以后刺激仍然在作用,但传入冲动频率可以逐渐降低到零;慢适应感受器以肌梭、颈动脉窦压力感受器为代表,它们在刺激持续作用时,一般只是在刺激开始以后不久出现一次冲动频率的某些下降,但以手可以较长时间维持在这一水平,直至刺激撤除为止。感受器适应的快慢各有其生理意义,如触觉的作用一般在于探索新异的物体或障碍物,它的快适应有利于感受器及中枢再接受新事物的刺激;慢适应感受器则有利于机体对某些功能状态如姿势、血压等进行长期持续的监测,有利于对它们可能出现的波动进行随时的调整。适应并非疲劳,因为对某一刺激产生适应之后,如增加此刺激的强度,又可以引起传入冲动的增加。
感受器产生适应的机制比较复杂,有的发生在刺激引起发生器电位这一阶段;有的发生在发生器电位诱发神经动作电位这一阶段。有不少感受器,适应发生的快慢一感受末稍所具有的附属结构有关。一个有趣的例子是,作为触压感受器的皮肤(如肠系膜)环层小体,其环层结构的存在与它适应有快速出现有关:实验中如果细心剥除环层结构后,直接轻压裸露的神经末稍仍可引起传入冲动发放,而且在这种情况下感受末稍变得不易适应,与剥除环层结构前表现的快适应明显不同。这个现象的解释是,当压力直接作用于环层结构表面时,压力要经过此结构才能传递到感受末稍表面,但因为环层结构具有一定的弹性,它受压后的弹性变形和回弹有可能使末稍表面实际受到的压力减轻或消失,使刺激的实际作用减弱以至全不起作用。
在人体的主观感受方面,也常常体验到类似“入芝兰室,久而不闻其香”之类的感觉适应现象。感觉适应的产生机制可能更为复杂,其中只部分地与感受器的适应有关,因为适应的产竹敢与传导途径中的突触传递和感觉中枢的某些功能改变有关。
第二节 视觉器官
引起视觉的外周感受器官是眼,它由含有感光细胞的视网膜和作为附属结构的折光系统等部分组成。人眼的适宜刺激是波长370-740nm的电磁波;在这个可见光谱的范围内,人脑通过接受来自视网膜的传入信息,可以分辨出视网膜像的不同亮度和色泽,因而可以看清视野内发光物体工反光物质的轮廓、形状、颜色、大小、远近和表面细节等情况。自然界形形色色的物体以及文字、图形等形象,通过视觉系统在人脑得到反映。据估计,在人脑获得的全部信息中,大约有95%以上来自视觉系统,因而眼无疑是人体最重要的感觉器官。
人眼的基本结构如图9-2所示。除了控制眼球运动的眼外肌和起保持、营养作用的巩膜、脉络膜等结构外,眼内与视觉传入信息的产生直接有关的功能结构,是位于眼球正中线上的折光系统和位于眼球后部的视网膜。由角膜经房水、晶状体、玻璃体直至视网膜的前表面,都是一些透明而无血管分布的组织,它们构成了眼内的折光系统,使来自眼外的光线发生折射,最后成像在视网膜上,视网膜具有同神经组织类似的复杂结构,其中包含有对光刺激高度敏感的视杆和视锥细胞,能将外界光刺激所包含的视觉信息转变成为电信号,并在视网膜内进行初步处理,最后以视神经纤维的动作电位的形式传向大脑。因此,形容眼的功能首先要研究眼内折光系统的不学特性,搞清楚它们怎样能把不同远近的物体成像在视网膜上以及形成清晰物像的限度;其次要阐明视网膜是怎样对视网膜成像进行换能和编码的。
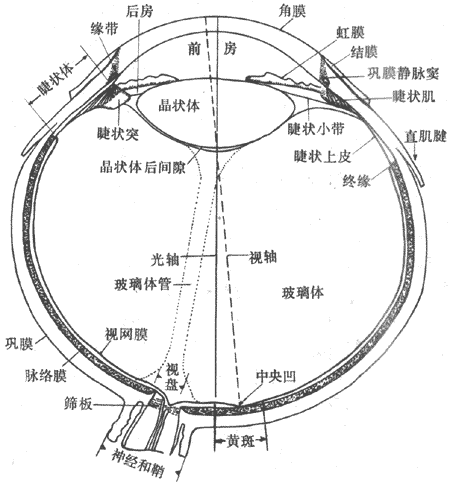
图9-2 眼球的水平切面(右眼)
一、眼的折光系统及其调节
当光线由 空气进入另一媒质构成的单球面折光体时,它进入物质的折射情况决定于该物质与空气界面的曲率半径R和该物质的折光指数n2;若空气的折光指数为n1,则关系式为
n2R/(n2-n1)=F2 (1)
F2称为后主焦距或第2焦距(空气侧的焦距为前主焦距或第一焦距),指由折射面到后主焦点的距离,可以表示这一折光的折光能力。表示折光体的折光能力还可用另一种方法,即把主焦距以m(米)作单位来表示,再取该数值的倒数,后者就称为该折光体的焦度(diopter);如某一透镜的主焦距为10cm,这相当于0,1m,则该透镜的折光能力为10焦度(10D)。通常规定凸透镜的焦度为正值,凹透镜的焦度为负值。
主焦距是一个折光体最重要的光学参数,由此可算出位于任何位置的物体所形成的折射像的位置。以薄透镜为例,如果物距α是已知的,像距b可由下式算出:
1/a+1/b=1/F2 (2)
由式(2)可以看出,当物距a趋于无限大时,1/a趋近于零,于是1/b接近于1/F2,亦即像距b差不多和F2相等;这就是说,当物体距一个凸透镜无限远时,它成像的位置将在后主焦点的位置。同样不难看出,凡物距小于无限大的物体,它的像距b恒大于F2,即它们将成像在比主焦点更远的地方。以上两点结论,对于理解眼的折光成像能力十分重要。
另外,根据光学原理,主焦点的位置是平行光线经过折射后聚焦成一点的位置,这一结论与上面提到的第一点结论相一致。每一物体的表面,都可认为是由无数的发光点或反光点组成,而由每一个点发出的光线都是辐散形的;只有这些点和相应的折射面的距离趋于无限大时,由这些点到达折射面的光线才能接近于平行,于是它们经折射后在主焦点所在的面上聚成一点,整个物质就达个面上形成物像。当然,无限过的概念本身决定了它是一个不可能到达的位置,实际上对人眼和一般光学系统来说,来自6m以外物体的各光点的光线,都可以认为是近于平行的,因而可能在主焦点所在的面上形成物像。
(二)眼的折光系统的光学特性
当用上述光学原理分析眼的折光特性时,首先遇到的一个困难是,眼球并非一个薄透镜或单球面折光体,而是由一系列由率半径和折光指数都不相同的折光体所组成的折光系统。显然,人眼折光系统的后主焦距不能简单地由式(1)算出,不过它的最主要的折射发生在角膜,而按几何学原理进行较复杂的计算,还是可以追踪出光线经眼内多个折光面行进的途径,并得出由这些组合的透镜组所决定的后主焦点的所在位置。
计算结果表明,正常成人眼处于安静而不进行调节的状态时,它的折光系统的后主焦点的位置,正好是其视风膜所在的位置。这一解剖关系对于理解正常眼的折光成像能力十分重要。它说明,凡是位于眼前方6m以外直至无限远处的物体,根据式(2)或由于由它们发出或反射出的光线在到达眼的折光系统时已近于平行,因而都可以在视网膜上形成基本清晰的像,这正如放置于照相机主焦点处的底片,可以拍出清晰的远景一样。当然,人眼不是无条件的看清任何远处的特体,例如,人眼可以看清楚月亮(或其他更远的星体)和它表面较大的阴影,但不能看清楚月球表面更小的物体或特征。造成后一限制的原因是,如果来自某物体的光线过弱,或它们在空间处女内传播时被散射或吸收,那么它们到达视网膜时已减弱到不足以兴奋感光细胞的程度,这样就不可能被感知;另外,如果物体过小或它们离眼的距离过大,则它们在视网膜上形成的大小,将会小到视网膜分辨能力的限度以下,因而也不能感知。
(三)眼的调节
如果安静状态的眼的折光能力正好把6m以外的物体成像在视网膜上,那么来自较6m为近的物体的光线将是不同程度呈辐射状的,它们在折射后的成像位置将在主焦点,亦即视网膜的位置之后;由于光线到达视网膜时尚未聚焦,因而物像是模糊的,由此也只能引起一个模糊的视觉形象。但正常眼在看近特时也十分清楚,这是由于眼在看近物时已进行了调节(accommodation),使进入眼内的光线经历较强的折射,结果也能成像在视网膜上。人眼的调节亦即折光能力的改变,主要是靠晶状体形状的改变;这是一个神经反射性活动,其过程如下:当模糊的视觉形象出现在视区皮层时,由此引起的下行冲动经锥体束中的皮层-中脑束到达中脑的正中核,再到达发出动眼神经中副交感节前纤维的有关核团,最后再经睫状神经节到达眼内睫状肌,使其中环行肌收缩,引起连接于水晶体囊的悬韧带放松;这样就促使水晶体由于其自身的弹性而向前方和后方凸出(以前突较为明显),使眼的总的折光能力较安静时增大,使较辐射的光线提前聚焦,也能成像在视网膜上。因9-3表示调节前后晶状体形状的改变。很明显,物体距眼球愈近,到达眼的光线辐散程度愈大,因而也需要晶状体作更大程度的变凸。调节反射进行时,除晶状体的变化外,同时还出现瞳孔的缩小和两眼视轴向鼻中线的会聚,前者的意义在于减少进入眼内光线的量(物体移近时将有较强光线到达眼球)和减少折光系统的球面像差和色像差;两眼会聚的意义在于看近物时物像仍可落在两眼视网膜的相称位置。
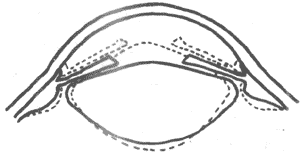
图9-3 眼调节前后睫状体位置和晶状体形状的改变
实线为安静时的情况,虚线为看近物经过调节后的情况,注意晶状体的前凸比后凸明显
人眼看近物的能力,亦即晶状体的调节能力是有一定限度的,这决定于水晶体变凸的最大限度。随着年龄的增加,水晶体自身的弹性将下降,因而调节能力也随年龄的增加而降低。眼的最大调节能力可用它所能看光天化日物体的最近距离来表示,这个距离或限度称为近点。近点愈近,说明晶状体的弹性愈好,亦即它的悬韧带放松时可以作较大程度的变凸,因而使距离更近的物体也能成像在视网膜上。例如,8岁左右的儿童的近点平均约8.6cm,20岁左右的成为约为10.4cm,而60岁时可增大到83.3cm。
(四)简化眼和视敏度
由于眼内有多个折光体,要用一般几何光学的原理画出光线在眼内的行进途径和成像情况时,显得十分复杂。因此,有人根据眼的实际光学特性,设计一些和正常眼在折光效果上相同、但更为简单的等效光学系统或模型,称为简化眼。简化眼只是一种假想的人工模型,但它的光学参数和其它特性与正常眼等值,故可用来分析眼的成像情况和进行其他计算。常用的一种简化眼模型,设想眼球由一个前后径为20mm的单球面折光体构成,折光指数为1.333;外界光线只在由空气进入球形界面时折射一次,此球面的曲率半径为5mm,亦即节点在球形界面后方5mm的位置,后主焦点正相当于此折光体的后极。显然,这相模型和正常安静的人眼一样,正好能使平行光线聚焦在视网膜上(图9-4)。
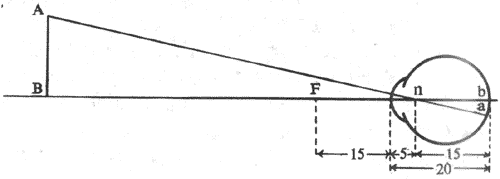
图9-4 简化眼及其成像情况
n为节点,AnB和anb是两个相似三角形;如果物距为已知,就可由物体大小算出物像
大小,也可算出两三角形对顶角(即视角)的大小
利用简化眼可以方便地计算出不远近的物体在视网膜上成像的大小。如图9-4所示,AnB和and是具有对顶角的两个相似的三角形,因而有:
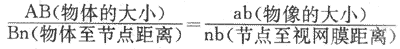
其中nb固定不变,相当于15mm,那么根据物体的大小和它距眼的距离,就可算出物像的大小。此外,利用简化眼可以算出正常人眼所能看清的物体的视网膜像大小的限度。检查证明,正常人眼即使在光照良好的情况下,如果视网膜小于5μm,一般就不能引起清晰的视觉。这说明,正常人的视力或视敏度(visualacuity)有一个限度;要表示这个限度,只能用人所能看清的最小视网膜的大小,而不能用所能看清的物体大小表示,因为物像有大小与物体的大小有关,大致相当于视网膜中央凹处一个视锥细胞的平均直径(但有些视锥的直径可小于2μm)。
通常用业检查视敏度的国际通用的视力表,就是近上述原理设计的。当人眼能看清5m处的一个圆形或E字形上相距1.5mm的缺口的方向时,按简化眼计算,此缺口在视网膜像中的距离约为5μm(实际计算值为 4.5μm),说明此眼视力正常,定为1.0;由图9-4也可以算出,当物像为5μm时,由光路形成的两个三角形的对顶角即视角约相当于1分度(即1');因此,如果受试者在视角为10分分度时才能看清相应增大了视力表上的标准图形的缺口(相当于国际视力表上最上面一排图),则视力定为0.1;在表上还列出视力0.2至0.9时的逐步减小的图形;但国际视力表上对这些相应图形的大小设计是有缺点的,如相当于0.2视力的图形比视力0.1的图形小1/2,而相当于视力1.0的图形只比视力为0.9时的图形小了1/9。这种表示视力方法显然不利于临床上表示视力的改善程度,例如由原来0.9的视力改善为1.0,或由0.1的视力改善为0.2,虽然视力都增加了0.1,但其真正改善的程度并不一样,因而不能作为统计处理的数据。为了避免这一缺点,我国有人设计了一种对数视力表(缪天荣,1966),它把国际视力表上记为1.0的正常视力记为5.0,而将视角为10分度时的视力记为4.0,其间相当于视力4.1、4.2直至4.9的图形,各比上一排形成的视角小=1.259……倍,而log=0.1;这样,视力表上不论原视力为何值,改善程度的数值都具有同样的意义。
眼的折光能力和调节能力异常 正常眼的折光系统在无需进行调节的情况下,就可使平行光线聚焦在视网膜上,因而可看清远处的物体;经过调节的眼,只要物体的距离不小于近点的距离,也能在视网膜上形成清晰的像被看清,此称为正视眼。若眼的折光能力异常,或眼球的形态异常,使平行光线不能在安静未调节的眼的视网膜上成像,则称为非正视眼,其中包括近视、远视和散光眼。有些眼静息时折光能力正常,但由于水晶体的弹性减弱或丧失,看远物时的调节能力减弱,此称为老视。
近视 多数由于眼球的前后径过长(轴性近视),致使来自远方物体的平行光线在视网膜前即已聚焦,此后光线又开始分散,到视网膜时形成扩散开的光点,以致物像模糊。便近视看近物时,因这时聚焦的位置较平行光线时为后,因而眼无需进行调节或进行较小程度的调节,就可在视网膜上成像;这就使近视能看清近物,且远点比正常眼还要近。纠正近视眼的方法是在眼前增加一个一定焦度的凹透镜片,使入眼的平行光线适当辐散,以便聚焦位置移后,正好能成像在视网膜上;这样使远物可以看清,而近物则像正常眼一样,依靠眼睛自身的调节能力。近视也可由于眼的折光能力超过正常,使平行光线成像在位置正常的视网膜之前,这种近视特称为屈光近视。
远视 由于眼球前后径过短,以致主焦点的位置实际在视网膜之后,这样入眼的平行光线在到达视网膜时尚未聚焦,也形成一个模糊的像,引起模糊的视觉。这时,患者在看远物时就需使自己的调节能力,使平行光线能提前聚焦,成像在位置前的视网膜上。由此可见,远视眼的特点是在看远物时即需动用眼的调节能力,因而看近物时晶状体的凸出差差不多已达到它的最大限度,故近点距离较正常人为大,视近物能力下降,纠正的方法是戴一适当焦度的凸透镜,使看远时不需晶状体的调节亦能在像在视网膜上,于是通过调节能力就可像正视眼一样用来看近物了。
散光 正常眼的折光系统的各折光面都是正球面的,即在球表面任何一点的曲率半径都是相等的。如果由于某些原因,折光面(通常见于角膜)在某一方位上曲率半径变小,而在与之相垂直的方位上曲率半径变大(相当于在一个硬的桌面上轻压一个乒乓球时,球面的曲率半径在垂直的方位上变小,在横的方位上变大一样),在这种情况下,通过角膜不同方位的光线在眼内不能同时聚焦,这会造成物像变形和视物不清。这种情况属于规则散光,可用适当的柱面镜纠正,后者的特点正是互相垂直方位上具有不同的曲率半径,当它和角膜的曲率半径改变大小相抵消时,使角膜的曲率异常得到纠正。
二、瞳孔和瞳孔对光反应
瞳孔指虹膜中间的开孔,是光线进入眼内的门户;它在亮光处缩小,在暗光处散大。虹膜由多单位平滑肌构成;在瞳孔周围的是环形肌层,受动眼神经中的副交感神经纤维支配,收缩时使瞳孔缩小,故又称瞳孔括约肌;虹膜的外周部分是辐散状肌纤维,受由颈部上行的交感神经纤维支配,收缩时使瞳孔散大,故又称瞳孔散大肌。瞳孔的大小可以控制进入眼内的光量。一般人瞳孔的直径可变动于1.5-8.0mm之间。假定人由光亮处进入暗室时瞳孔直径可增加5倍,那么瞳孔的受光面积应增大25倍;可见瞳孔的变化,有保持在不同光照情况下进入眼内的光量较为恒定的作用。但暗室中较强阳光照射的光照强度实际减弱约100万倍,因而单靠瞳孔大小的改变,远不足以使进入眼内的光量保持恒定。事实上,人眼在不同的亮度情况下是靠视网膜中不同的感光细胞来接受光刺激的,在暗光处起作用的视杆细胞对光的敏感程度要比在亮光处起作用的视锥细胞大得多,因此在暗处看物,只需进入眼内光量适当增加即可。由此可见,通过改变瞳孔大小以调节进入眼内的光量还是有一定意义的。
瞳孔大小随光照强度而变化的反应,是一种神经反射,称为瞳孔对光反射。引起此反射的感受器就是视网膜,传入纤维在视神经中,但这部分纤维在进入中枢后不到达外侧膝状体,而在在中脑的顶盖前区换神经元,然后到同侧和对侧的动眼神经核,传出纤维主要是动眼神经中的副交感纤维,效应器也主要是瞳孔约肌。
瞳孔对光反应的特点是效应的双侧性,即如果光照的是一侧眼睛时,除被照眼出现瞳孔缩小外,同时未受光照拐殊途同归瞳孔也缩小,后者我为互感性对光反射。临床上有时可见到瞳孔对光反应消失、瞳孔左右不等、互感性瞳孔反应消失等异常情况,常常是由于与这些反射有关的反射绵弧某一部分受损的结果,因而可以藉瞳孔反应的异常帮助进行神经病变的定位诊断。
房水和眼内压 房水指充盈于眼的前、后房中的液体,其成分类似血浆,但蛋白质含量较血浆低得多,而HCO3-含量却超过血浆;因而房水的总渗透压也较血浆为高。房水的生成部位在睫状体脉络膜丛,生成后由后房经瞳孔进入前房,再由巩膜和角膜结合处的前房角进入巩膜静脉窦,最后汇入静脉系统。房水不断生成,不断回收入静脉,使它在后房和前房之间流动不息。据测定,正常时房水的生成速度每分钟约为2mm3;由于它的生成和回收之间保持着动态平衡,使得眼内政党时的房水量保持恒定,又由于眼的前、后房容积也是相对恒定的,于是使其中静水压(即眼内压)也保持相对的稳定。据国内调查资料,我国成年人眼内压正常值为2.27-3.2kPa(17-24mmHg),平均2.67kPa(17-24mmHg)。
眼内压的相对稳定,对保持眼球特别是角膜的正常开头和折光能力有重要的意义。当眼球被刺穿时,可能导致房水流失,眼内压下降,引起眼球变形,角膜也不能保持正常的曲度。人眼的总折光能力与眼内折光体都有一定关系,但最主要的折射发生在空气与角膜接触的界面上。这约占总折光能力的80%。因此,角膜的曲度和形状的改变将显著地影响眼的折光能力,严重地影响视力。房水也对它所接触的无血管组织如角膜和晶状体起着营养的作用。房水循环障碍时会造成眼内压过高,临床上称为青光眼,可导致角膜、晶状体以及虹膜等结构的代谢障碍,严重时造成角膜混浊、视力丧失。
房水生成的机制目前尚不完全明了。通常认为,除了在睫状体脉络膜丛处的毛细血管靠被动滤过(类似在一般毛细血管的动脉端生成组织液,见第四章),使血浆中的水分和盐类透出血管壁生成房水外,还有主动过程的参与,否则就难于解释房水何以有较血浆中浓度高的HCO3-等盐类离子。用组织化学的方法证明,睫状体上皮细胞含有较多的碳酸酐酶,此酶的作用是使细胞代谢过程中产生的CO2和H2O,迅速生成H2CO3,并解离出HCO3-,后者经膜上的主动转运过程进入房水,造成它的房水中的高浓度,这个高浓度造成的负电位和高渗透压还能进一步促使血浆中的Na+和水分子进入房水。临床上可以使用碳酸酐酶抑制剂(如乙酰唑胺)减少房水生成,降低眼内压,其作用机制与上述的房水生成机制有关。
三、视网膜的结构和两种感光换能系统
来自外界物体的光线,通过眼内的折光系统在视网膜上形成物像,是视网膜内的感光细胞被刺激的前提条件。视网膜像还有一个物理范畴内的内像,用几何光学的原理可以较容易地对它加以说明,和外界物体通过照相机的中的透镜组在底片上形成的物像并无原则上的区别;但视觉系统最后在主观意识上形成的“像”,则是属于意识或心理范畴的主观印象,它由来自视网膜的神经信息最终在大脑皮层等中枢结构内形成。作为感受器生理,重点是视网膜怎样把物理像转换成视神经纤维上的神经信号,以及在这些信号的序列和组合中怎样包括了视网膜像、亦即外界物体所提供的信息内容。应该提出,视觉研究的进展虽然较快,但也只是初步的。
(一)视网膜的结构特点
视网膜的厚度只有0.1-0.5mm,但结构十分复杂。它的主要部分在个体发生上来自前脑泡,故属于神经性结构,其中细胞通过突触相互联系。经典组织学将视网膜分为十层,但按主要的细胞层次简化为四层业描述,如图9-5所示。从靠近脉络膜的一侧算起,视网膜最外层是色素细胞层;这一层的来源不属神经组织,血液供应也来自脉络膜一侧,与视网膜其他层接受来自视网膜内表面的血液供应有所不同;临床上见到的视网膜剥离,就发生在此层与其它层次之间。色素细胞层对视觉的引起并非无关重要,它含在黑色素颗粒和维生素A,对同它相邻接的感光细胞起着营养和保护作用。保护作用是除了色素层可以遮继来自巩膜侧的散射光线外,色素细胞在强光照射视网膜时可以伸出伪足样突起,包被视杆细胞外段,使其相互隔离,少受其他来源的光刺激;只有在暗光条件下,视杆外段才被暴露;色素上皮的这种活动受膜上的多巴胺受体控制。此层内侧为感光细胞层。在人类和大多数哺乳动作动物,感光细胞分视杆和视锥细胞两种,它们都含有特殊的感光色素,是真正的光感受器细胞。视杆和视锥细胞在形态上都可分为四部分,由外向内依次称为外段、内段、胞体和终足(图9-6);其中外段是感光色素集中的部位,在感光换能中起重要作用。视杆和视锥细胞在形成上的区别,也主要在外段它们外形不同,所含感光色素也不同。视杆细胞外段呈长杆状,视锥细胞外段呈圆锥状。两种感光细胞都通过终足和双极细胞层内的双极细胞发生突触联系,双极细胞一般再和节细胞层中的神经节细胞联系。视网膜中除了这种纵向的细胞间联系外,还存在横向的联系,如在感光细胞层和双极细胞层之间有水平细胞,大双极细胞层和节细胞层之间有无长突细胞;这些细胞的突起在两层细胞之间横向伸展,可以在水平方向传递信息,使视网膜在不同区域之间有可能相互影响;这些无长突细胞还可直接向节细胞传递信号。近年来发现,在视网膜还存在一种网间细胞,它的细胞体位于双极细胞层和节细胞层之间,但突起却伸到感光细胞层和双极细胞层。如果把感光细胞经过双极细胞到神经节细胞的途径,看作是视觉信息的初始阶段。近年来还发现,视网膜中除了有通常的化学性突触外,还有大量电突触存在。由此可见,视网膜也和神经组织一样,各级细胞之间存在着复杂的联系,视觉信息最初在感光细胞层换能变成电信号后,将在视网膜复杂的神经元网络中经历某种处理和改变,当视神经纤维的动作电位序列作为视网膜的最终输出信号传向中枢时,它们已经是经过初步加工和处理的信息了。
盲点 由节细胞层发出的神经轴突,先在视网膜表面聚合成一整束,然后它透视网膜,在眼的后极出眼球,这就在视网膜表面形成视神经乳头。在乳头的范围内,实际上没有视网膜特有的细胞结构,因而落于该处的光线或视网膜像的组成部分,将不可能被感知,故称为盲点。两侧视神经乳头在视网膜内黄斑或中央凹中心的鼻侧约3mm处。但正常时由于用两眼看物,一侧盲点可以被对侧视觉补偿,人们并不觉察自己的视野中有一处无视觉感受的区域。盲点的存在可用专门设计的方法来证明。
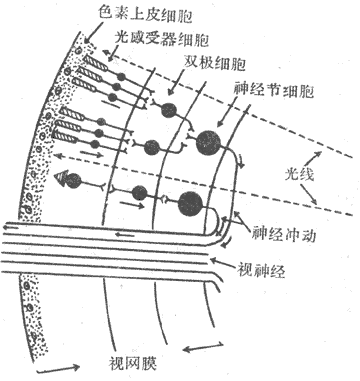
图9-5 视网膜的主要细胞层次及其联系模式图
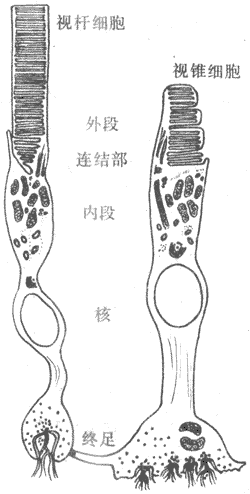
图9-6 哺乳动物光感受器细胞模式图
(二)视网膜的两种感光换能系统
根据对视网膜结构和功能的研究,目前认为在人和大多数脊椎动物的视网膜中存在着两种感光换能系统。一种由视杆细胞和与它们相联系的双极细胞和神经节细胞等成分组成,它们对光的敏感度较高,能在昏暗的环境中感受光刺激而引起视觉,但视物无色觉而只能区别明暗;且视物时只能有较粗略的轮廓,精确性差,这称为视杆系统或晚光觉系统;另一种由视锥细胞和与它们有关的传递细胞等成分组成,它们对光的敏感性较差,只有在类似白昼的强光条例下才能被刺激,但视物时可辨别颜色,且对物体表面的细节和轮廓境界都能看得很清楚,有高分辨能力,这称为视锥系统或昼光觉系统(前述视敏度的测定实际是视锥系统视力的测定)。
证明这两种相对独立的感光-换能系统存在的主要依据是:①人视网膜中视杆和视锥细胞在空间上的分布是不均匀的,愈近视网膜周边部,视杆细胞愈多而视锥细胞愈少;愈近视网膜中心部,视杆细胞愈少而视锥细胞愈多;在黄斑中心的中央凹处,感光细胞全部是视锥而无视杆细胞;与上述细胞分布相对应,人眼视觉的特点正是中央凹在亮光处有最高的视敏度和色觉,在暗外则中央视力较差;相反地,视网膜周边部则能感受弱光的刺激,但这时无色觉而清晰度较差。②两种感光细胞和双极细胞以及节细胞形成信息传递通路时,逐级之间都有一定程度的会聚现象(参看第十章),但这种会聚在视锥系统程度较小,在中央凹处甚至可以看到一个视锥细胞只同一个双极细胞联系,而这个双极细胞也只同一个神经节细胞联系的情况,这种低程度会聚或无会聚现象的“单线联系”,显然是视锥系统有较高的精细分辨能力的结构基础;与此相对照,在视杆系统则普遍存在多个感光细胞同一个双极细胞联系,而多个双极细胞再同一个神经节细胞联系的会聚式排列;在视网膜周边部,可看到多达250个视杆细胞经少数几个双极细胞会聚于一个神经节细胞的情况;在这种情况下,当然不能期待这样的感觉系统有高的精细分辨能力。但这样的聚合系统却具有较强的总和多个弱刺激的能力。③从动物各系统特点来看,某些只在白昼活动的动物如爬虫类和鸡等,视网膜全无视杆而只胡视锥细胞。而另一些只在夜间活动的动物如地松鼠和猫头鹰等,视网膜中只胡视杆而不含视锥细胞。④视杆细胞中只含有一种感光色素,即视紫红质(rhodopsin),而视锥细胞却因所含感光色素的吸收光谱特性不同而分为三种,这是同视杆系统无色觉而视锥系统有色觉的事实相一致的。
四、视杆细胞的感光换能机制
从上世纪末开始,有人就从视网膜中提取出了一定纯度的感光色素即视紫红质,它在暗处呈红色;实验中还可以证明,提取出来的这种感光色素对不同波长光线的吸收光谱,基本上和晚光觉对光谱不同部分的敏感性曲线相一致(图9-7)。这一事实十分重要,因为既然光线对某种感光色素的光化学作用的强度正好与这些光线所引起的视觉的强度相一致,那就是提示前者可能是后者的基础。
(一)视紫红质的光化学反应及其代谢
视紫红质的分子量约为27-28kd,是一种与结合蛋白质,由一分子称为视蛋白(opsin)的蛋白质和一分子称为视黄醛(retnal)的生色基团所组成。视蛋白的肽链序列已搞清,它的肽链中有7段穿越所在膜结构、主要由疏水性氨基酸组成的α-螺旋区段,同一般的细胞膜受体具有类似的结构。视黄醛由维生素A变来,后者是一种不饱和醇,在体内一种酶的作用下可氧化成视黄醛。提纯的视紫红质在溶液中对500nm波长的光线吸收能力最强,这与人眼在弱光条件下对光就业上蓝绿光区域(相当于500nm波长附近)感觉最明亮(不是感到了蓝绿色)的事实相一致(图9-7),说明人在暗视觉与视杆细胞中所含视紫红质的光化学反应有直接的关系。
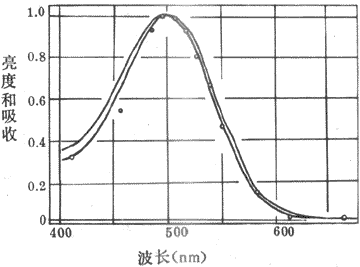
图9-7弱光条件下人眼所感到的光谱亮度曲线和实验条件下
视紫红质对光谱不同部分的吸收曲线视觉中最明亮的区域和视紫红制裁
吸收能力最强的部分都在500nm的波长附近
视紫红质在光照时迅速分解为视蛋白和视黄醛,这是一个多阶段的反应。目前认为,分解的出现首先是由于视黄醛分子在光照时发生了分子构象的改变,即它在视紫红质分子中本来呈11-顺型(一种较为弯曲的构象),但在光照时变为全反型(一种较为直的分子构象)。视黄醛分子构象的这种改变,将导致视蛋白分子构象也发生改变,经过较复杂的信号传递系统的活动,诱发视杆细胞出现感受器电位。据计算,一个光量子被视紫红质吸收,就足以使视黄醛分子结构发生改变,导致视紫红质最后分解为视蛋白和视黄醛。视紫红质分解的某些阶段伴有能量的释放,但这看来不是诱发感受器电位的直接原因。
在亮处分解的视紫红质,在暗处又可重新合成,亦即它是一个可逆反应,其反应的平衡点决定于光照的强度。视紫红质再合成的第一步,是全反型的视黄醛变为11-顺型的视黄醛,很快再同视蛋白结合。此外,贮存在视网膜的色素细胞层中的维生素A也是全反型的,它们也可在耗能的情况下变成11-顺型的,进入视杆细胞,然后再氧化成11-顺型的视黄醛,参与视紫红质的合成补充;但这个过程进行的速度较慢,不是促进视紫红制裁再合成的即时因素。人在暗处视物时,实际是既有视紫红质的分解,又有它的合成,这是人在暗光处能不断视物的基础;光线愈暗,全盛过程愈超过分解过程,视网膜中处于合成状态的视紫红质数量也愈高,这也使视网膜对弱光愈敏感;相反,人在亮光处时,视紫红质的分解增强,合成过程甚弱,这就使视网膜中有较多的视紫红质处于分解状态,使视杆细胞几乎失去了感受光刺激的能力;事实上,人的视觉在亮光处是靠另一种对光刺激较不敏感的感光系统即视锥来完成的,后一系统在弱光时不足以被刺激,而在强光系统下视杆细胞中的视紫红质较多地处于分解状态时,视锥系统就代之而成为强光刺激的感受系统。在视紫红质和再合成的过程中,有一部分视黄醛被消耗,这最终要靠由食物进入血液循环(相当部分贮存于肝)中的维生素A来补充。长期摄入维生素A不足,将会影响人在暗光处的视力,引起夜盲症。
(二)视杆细胞外段的超威结构和感受器电位的产生
感光细胞的外段是进行光-电转换的关键部位。视杆细胞外段具有特殊的超威结构,如图9-8所示。在外段部分,膜内的细胞浆甚少,绝大部分为一些整齐的重叠成层的圆盘状结构所占据,这圆盘称为视盘。每一个视盘是一个扁平的囊状物,囊膜的结构和细胞膜类似,具有一般的脂质双分子层结构,但其中镶嵌着的蛋白质绝大部分是视紫红质,亦即视杆细胞所含的视紫红质实际上几乎全部集中在视盘膜中。视盘的数目在不同动物的视杆细胞中相差很大,人的每个视杆细胞外段中它们的数目近千;每一个视盘所含的视紫红质分子约有100万个。这样的结构显然有利于使进入视网膜的光量子有更大的机会在外段中碰到视紫红质分子。
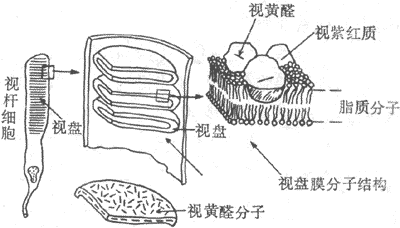
图9-8 视杆细胞外段的超微结构示意图
有人用细胞内微电极技术,研究了视杆细胞外段内外的电位差在光照前后的变化,结果发现在视网膜未经照射时,视杆细胞的静息电位只有-30?/FONT>-40mV,比一般细胞小得多。经分析表明,这是由于外段膜在无光照时,就有相当数量的Na+通道处于开放状态并有持续的Na+内流所造成,而内段膜有Na+泵的连续活动将Na+移出膜外,这样就维持了膜内外的Na+平衡。当视网膜受到光照时,可看到外段膜两侧电位短暂地向超极化的方向变化,由此可见,外段膜同一般的细胞膜不一致,它是在暗处或无光照时处于去极化状态,而在受到光刺激时,跨膜电痊反而向超极化方向变化,因此视杆细胞的感受器电位(视锥细胞也一样),表现为一种超极化型的慢电位,这在所有被研究过的发生器或感受器电位中是特殊的,它们一般都表现为膜的暂时去极化。
光子的吸收引起外段膜出现超极化电反应的机制已基本搞清,这就是光量子被作为受体的视紫红质吸收后引起视蛋白分子的变构,又激海参了视盘膜中一种称为传递蛋白(transducin)Ct的中介物,后者在结构上属于G-蛋白家庭的一员,它激活的结果是进而激活附近的磷酸二酯酶,于是使外段部分胞浆中的cGMP大量分解,而胞浆中cGMP的分解,就使未受光刺激时结合于外段膜的cGMP由也膜解离而被分解,而cGMP在膜上的存在正是这膜中存在的化学门控式Na+通道开放的条件,膜上cGMP减少,Na+通道开放减少,于是光照的结果出现了我们记录到的超极化型感受器电位。据估计,一个视紫红质被激活时,可使约500个传递蛋白被激活;虽然传递蛋白激活磷酸二酯酶是1对1的,但一个激活了的磷酸二酯酶在一秒钟内大约可使4千多个cGMP分子降解。由于酶系统的这种生物放大作用,就可以说明1个光量子的作用何以能在外段膜上引起大量化学门控式Na+通道的关闭,引起一个足以为人的视觉系统所感知的超极化型电变化。
视杆细胞外段和整个视杆细胞都没有产生动作电位的能力,由光刺激在外段膜上引起的感受器电位只能以电紧张性的扩布到达它的终足部分,影响终点(相当于轴突末稍)外的递质释放。
五、视锥系统的换能和颜色视觉
视锥系统外段也具有与视杆细胞类似的盘状结构,并含有特殊的感光色素,但分子数目较少。已知,大多数脊椎动物具有三种不同的视锥色素,各存在于不同的视锥细胞中。三种视锥色素都含有同样的11-顺型视黄醛,只是视蛋白的分子结构稍有不同。看来是视蛋白分子结构中的微小差异,决定了同它结合在一起的视黄醛分子对何种波长的光线最为敏感,因而才有视杆细胞中的视紫红质和三种不同的视锥色素的区别。光线作用于视锥细胞外段时,在它们的外段膜两侧也发生现视杆细胞类似的超级化型感受器电位,作为光-电转换的第一步。目前认为视锥细胞外段的换能机制,也与视杆细胞类似。
视锥细胞功能的重要特点,是它有辨别颜色的能力。颜色视觉是一种复杂的物理-心理现象,颜色的不同,主要是不同波长的光线作用于视网膜后在人脑引起的主观印象。人眼一般可在光谱上区分出红、橙、黄、绿、青、蓝、紫等七种颜色,每种颜色都与一定波长的光线相对应;但仔细的检查可以发现,单是人眼在光谱可区分的色泽实际不下150种,说明在可见光谱的范围内波长长度只要有3-5nm的增减,就可被视觉系统分辨为不同的颜色。很明显,设想在视网膜中存在上百种对不同波长的光线起反应的视锥细胞或感光色素,是不大可能的。但物理学上从牛顿的时代或更早就知道,一种颜色不仅可能由某一固定波长的光线所引起,而且可以由两种或更多种其他波长光线的混合作用而引起。例如,把光谱上的七色光在所谓牛顿色盘上旋转,可以在人眼引起白色的感觉;用红、绿、蓝三种色光(不是这三种颜色的颜料)作适当混合,可以引起光谱上所有任何颜色的感觉。这后一现象特别重要;这种所谓三原色混合原理不仅早已广泛地应用于彩色照像、彩色电视等方面,而且被用于说明颜色视觉的产生原理本身。早在上世纪初,Young(1809)和Helmholtz(1824)就提出了视觉的三原色学说,设想在视网膜中存在着分别对红、红、蓝的光线特别敏感的三种视锥细胞或相应的三种感光色素,并且设想当光谱上波和介于这三者之间的光线作用于视网膜时,这些光线可对敏感波长与之相近两种视锥细胞或感光色素起不同程度的刺激作用,于是在中枢引起介于此二原色之间的其他颜色的感觉。视觉三原色学说用较简单的生物感受结构的假设说明了复杂的色觉现象,一般为多数人所接受;但在实验中试图寻找出游同种类的视锥细胞或感光色素长时间未获成功。用光学显微镜和电子显微镜不能发现视锥细胞之间在结构上有什么不同,同时也未能用一般的化学方法分离邮不同的视锥感光色素。
70年代以来,由于实验技术的进步,关于视网膜中有三种对不同波长光线特别敏感的视锥细胞的假说,已经被许多出色的实验所证实,例如,有人用不超过单个视锥直径的细小单色光束,逐个检查并绘制在体(最初实验是在金公和蝾螈等动物进行,以后是人)视锥细胞的光谱吸收曲线,发现所有绘制出来的曲线不外三种类型,分别代表了三类光谱吸收特性不同的视锥细胞,一类的吸收峰值在420nm外,一类在531nm外,一类在558nm外,差不多正好相当于蓝、绿、红三色光的波长,和上述视觉三原色学说的假设相符。用微电极记录单个视锥细胞感受器电位的方法,也得到了类似的结果,即不同单分光引起的超极化型感受器电位的大小,在不同视锥细胞是不一样的(图9-9),峰值出现的情况符合于三原色学说。
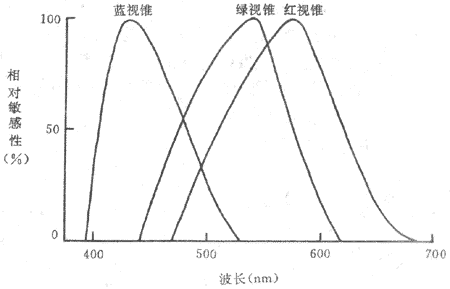
图9-9 人视网膜中三种不同视锥细胞的光谱相对敏感性
三原色学说和它的实验依据,大体上可以说明临床上遇到的所谓色盲和色弱的可能发病机制。红色盲也称第一色盲,被认为是由于缺乏对较长波长光线敏感的视锥细胞所致;此外还有绿色盲,也称第二色盲,蓝色盲也称第三原色盲,都可能是由于缺乏相应的特殊视锥细胞所致。红色盲和绿色盲较为多见,在临床上都不加以区别地称为红绿色盲;蓝色盲则极少见。色盲患者的颜色不仅不能识别绿色,也不能区分红也绿之间、绿与蓝之间的颜色等。有些色觉异常的人,只是对某种颜色的识别能力差一些,亦即他们不是由于缺乏某种视锥细胞,而只是后者的反应能力较正常人为弱的结果,这种情况有别于真正的色盲,称为色弱。色盲除了极少数可以由于视网膜后天病变引起外,绝大多数是由遗传因素决定的。
三原色学说虽然比较圆满地说明许多色觉现象和色盲产生的原因,并已在光感受细胞的一级得到了实验证实,但并不能解释所有的颜色视觉现象,如颜色对比现象就是一个例子。试将蓝色的小纸块放在黄色或其它颜色的背景上,会觉得放在黄色背景上那个蓝纸块特别蓝,同时觉得背景也比未放蓝纸块时更黄(在我国北方的黄土高原,当春天的风造成黄尘蔽日的情况时,会觉得平常的日晃灯管的光线变得较蓝了)。这种现象称为颜色对比,而黄和蓝则称为对比色或互补色。颜色对比现象只出现对比色之间,而不是任意的两种颜色之间。互为对比色的颜色对尚有:红一绿以及黑和白。根据颜色对比等不容易用三原色学说圆满视觉现象,几乎是在三原色学说提出的同时就出现了另一种色觉学说,称为对比色学说(Hering,1876)。该学说提出在视网膜中存在着三种物质,各对一组对比色的刺激起性质相反的反应。如前所述,近年来在视锥细胞一级进行的研究有利于三原色学说而不利于对比色学说,但后来在视网膜其它层细胞进行的一些实验却又符合对比色学说的推测。如在金鱼水平细胞进行的微电极研究说明,此类细胞和视杆、视锥细胞不同,既能出现超极化的跨膜电位改变,也能出现去极化型的电位改变,而且在用多种不同色光刺激时发现,有些水平细胞在黄光刺激时出现最大的去极化反应,在蓝光刺激时出出现最大的超极化型反应;另一些水平细胞则在红和绿色刺激时有类似的不同反应。这些现象是同对比色学说一致的。看来可能的是,各以部分色觉现象为出发点的两种色觉学说都是部分正确的,在视锥细胞一级,不同色光以引起三种不同视锥细胞产生不同大小的超极化型电变化进行编码;但到了水平细胞一级或其它级细胞(包括某些中枢神经元),信息又进行重新编码,不同颜色双可以用同细胞对互为对比色的颜色出现相反形式的电反应来编码。以上事实说明,颜色视觉的引起是一个十分复杂的过程,它需要有从视网膜视锥细胞到皮层神经元的多级神经成分的参与才能完成。
六、视网膜的信息处理
由视杆和视锥细胞产生的电信号,在视网膜内要经过复杂的细胞网络的传递,最后才能由神经节细胞发生的神经纤维以动作电位的形式传向中枢。由于视网膜内各种细胞之间的排列和联系非常复杂,与细胞间信息传递有关的化学物质种类繁多(除一般神经系统中常见的递质外,连同视网膜中已发现的各神经肽类物质,总数已达30余种),因而视觉信息在从感光细胞向节细胞传递时,必然要经历种种改变;这实际就是视网膜本身对视觉信息的初步处理,它是在视网膜特定的细胞构筑和化学构筑的网络中按照某些规律进行的,但对这些规律的了解还是很肤浅的。现在所能初步肯定的是,双极细胞、水平细胞和多数无长突细胞也同两种感光细胞一样,没有产生动作电位的能力(但部分无长突细胞可产生动作电位);但这三种细胞同感光细胞不同的是,它们在前一级细胞的影响之下,既能产生超级化型慢电位,也能产生去极化型慢电位(相当于一般神经元突触后膜处的IPSp 和EPSP,见第十章)。所有这些慢电位,只能作电紧张性的扩布(参看第二章局部兴奋),影响突触前膜递质释放量的改变,从而引起下一级细胞产生慢电位变化(也包括电突触性相互影响);只有当这样的慢电位传递到神经节细胞体时,由于后者有产生动作电位的能力,当两种形式的慢电位总和的结果,使节细胞的静息膜电位能够去极经到阈电位水平时,才会产生“全或无”式的动作电位,作为视网膜的最后输出信号传向中枢(图9-10)。
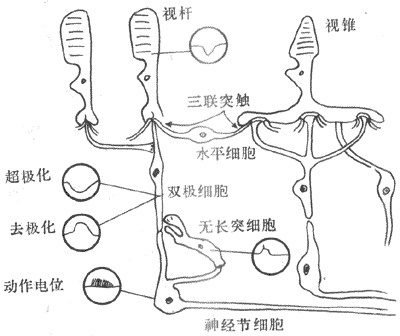
图9-10视网膜各类细胞排列及其产生的电反应的类型示意图
只有神经节细胞能产生动作电位
虽然关于视网膜中信息处理的细节还不很清楚,但不妨先把视网膜作为控制论中的“黑箱”来看待,只对它的输入(相当于作用于视网膜的光刺激)和它的输入(相当于由视神经传出的动作电位序列)进行对照分析,这样也可以初步理解视网膜信息处理和编码的最终结果。首先要记信一个事实是,视神经中纤维的总数(亦即节细胞的总数),只有全部感光细胞的1%。这一简单事实就足以说明,视神经不可能通过其纤维“点对点”地传递视网膜中各感光细胞被光照的情况(中央凹处少数视锥细胞例外);因而大多数视神经纤维所传递的信号,只能是决定于多个感光细胞并因而含较多的信息量。
用小的光点刺激猫视网膜并同时记录单条视神经纤维动作电位的方法表明,发出视神经纤维的节细胞大致可分为三类,分别称为X-、Y-和W-细胞。X-和Y-细胞的特点是它们都具有大致呈同心圆式的“中心-周边感受野”;一个节细胞的感受野在这里指视网膜上某一特定的区域,当后者受到刺激时可能使该节细胞发生反应;但这两种细胞的感受野都由两部分构成;当光线作用于感受野的中心部分时出现节细胞放电增加,而当光线作用于环绕该中心的一定范围内视网膜部分时,反而使该节细胞放电减少(图9-11,上),这是一种类型的中心-周边感受野,这就是当感受野的中心部分接受光刺激时引起节细胞放电数减少,而刺激其周边部分时,反而引起该细胞放电增多,这种节细胞可称为中心撤光反应细胞(图9-11,下)。
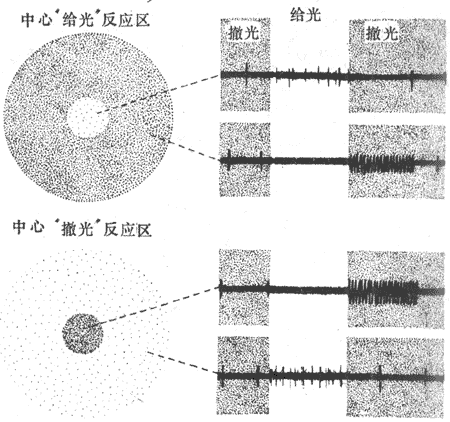
图9-11 视神经纤维的感受野和放电特征
X-和Y-细胞都可以有上述两种类型的感受野。它们之间的区别是,X-细胞的感受野较小,对刺激的反应是持续的;Y-细胞的感受野较大,而反应是时相性的,与刺激呈非线性关系。在猴子的实验中还发现,不同的X-细胞对不同波长的光线反应不同,但对光的强度变化不敏感;而Y-细胞则对光照的强度变化敏感,却对光线的波长变化不敏感。至于W-细胞,它们有较前两者大得多的感受野,或是在光刺激时有放电,或是在撤光时有放电,但对刺激的反应比较迟缓,一般没有性质相反的周边视野区域。从形态学上看,X-和X-细胞主要由双极细胞接受输入,而W细胞则主要接受无长突细胞的传入。
上述简单的事实说明,每一条视神经纤维上的神经冲动并不简单地表示某一部分视网膜受到光照或无光照,以上述X-为例,它的最强烈放电是出现在其感受野中心部分受到光照、而其外周部分全无光照(或相反的情况)的时候;如果中心和外周部同时受到光照,此节细胞的放电反而无大变化或只少有增加。另外,有事实说明,视网膜像经视网膜处理后,已被分解为不同“像素”,如有节细胞贡中枢传输图像组成波长的住处有的传输它的不同亮度。这种把感受信息分解为其组成的“要素”,再在感受通路中进行“平行”传输和处理,在脑的各种感觉功能的研究中经常遇到,但是人的视觉也和另的感觉一样,最复杂的信息处理和加工发生在中枢、特别是它的高级部分。
七、与视觉有关的其它现象
(一)暗适应和明适应
人从亮处进入暗室时,最初看不清楚任何东西,经过一定时间,视觉敏感度才逐渐增主,恢复了在暗处的视力,这称为暗适应。相反,从暗处初来到亮光处,最初感到一片耀眼的光亮,不能看清物体,只有稍待片刻才能恢复视觉,这称为明适应。
暗适应是人眼对光的敏感度在暗光处逐渐提高的过程。在进入暗室后的不同时间,连续测定人的视觉阈值,亦即测定人眼刚能感知的光刺激强度,可以看到此阈逐渐变小、亦即视觉的敏感度在暗处逐渐提高的过程。如图9-12所示,一般是在进入暗室后的最初约7分钟内,有一个阈值的明显下降期,以后又出现阈值的量明显下降;于进入暗室后的大约25-30分钟时,阈值下降到最低点,并稳定于这一状态。暗适应的产生机制与视网膜中感光色素在暗处时再合成增加,因而增加了视网膜中处于未分解状态的色素的量有关。据分析,暗适应的第一阶段主要与视锥细胞色素的合成量增加相一致;第二阶段亦即暗适应的主要构成部分,则与视杆细胞中视紫红质的合成增强有关。
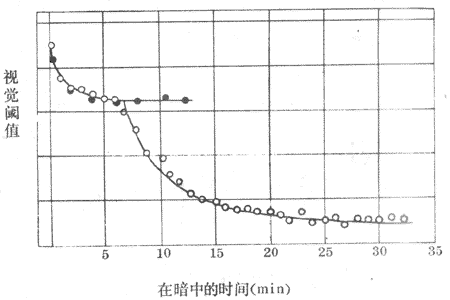
图9-12 暗适应曲线
。表示用白光对全眼的测定结果
·表示用红光对中央凹测定的结果(表示视锥细胞单独的暗适应曲线,
因中央凹为视锥细胞集中部位,且红光不易被视杆细胞所感受)
明适应出现较快,约需一分钟即可完成。耀眼的光感主要是由于在暗处蓄积起来的合成状态的视紫红质在进入亮处时先迅速分解,因为它对光的敏感性较视锥细胞中的感光色素为高;只有在较多的视杆细胞色素迅速分解之后,对光较不敏感的视锥细胞色素才能在亮光环境中感光。
(二)视野
单眼固定地注视前方一点不动,这时该眼所能看到的范围称为视野。视野的最大界限应以它和视轴(单眼注视外界某一点时,此点的像正好在视网膜黄斑中央凹处,连接这两点的假想线即视轴)所成夹角的大小来表示。在同一光照条件下,用不同颜色的目标物测得的视野大小不一样,白色视野最大,其次为黄蓝色,再次为红色,而以绿色视野为最小。设想视野的大小除与各类感光细胞在视网膜中的分布范围有关外,在细节上还缺乏圆满的解释。另外,由于面部结构阻挡视线,也影响视野的形状,如一般人颞侧视野较大,鼻侧视野较小等。临床医生检查视野,使用特制的视野仪,并用不同颜色的视标进行检查,目的在于了解视网膜的普遍感光能力,有时可藉以发现较大范围的视网膜病变。某些视网膜、视神经或视觉传导路的病变,有特殊形式的视野缺损,在诊断有意义。
(三)视网膜电图
将一个引导电极和角膜接触,另一个电极置于额部作为参考电极,当给视网膜以广泛光刺激时,可以在灵敏的电测量仪器上记录到一系列电变化,这称为视网膜电图(electroretinogram)。视网膜电图在性质上不同于同微电极在单一视网膜细胞成分记录到的电现象;视网膜电图是整个视网膜中各种成分在受到大范围光照时的多种电反应的综合反映。它通常由命名为a、b和c的三个波组成(图9-13)。据实验分析,a波主要来源于感光细胞的感受器电位;b波幅度较大,主要与双极细胞等细胞的活动有关;c波平缓而持续时间长,可能与色素细胞层的活动有关。有时在光照撤除时还可在缓慢持续的c波上再出现一个波动,称为d波,产生原因尚不明了。视网膜电图虽易于测量和描记,但反映视网膜功能状态或病变的特异性不大,目前只发现少数疾病时有特殊的视网膜电图改变,故在临床上作用的意义不很大。
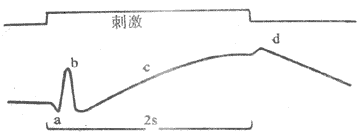
图9-13 猫的视网膜电图上面直线上的标记是给光和撤光时间
(四)双眼视觉和立体视觉
人和高等哺乳动物的双眼都在面部前方,视物时两眼视野差不多部分的像又各循自己特有的神经通路传向中枢,但正常人主观察感觉上只产生一个“物”的感觉。两眼视物而只产生一个视觉形象的前提条件是:由物质同一部分的的光线,应成像在两侧视网膜的相称点上。例如,两眼的黄斑部就互为相称点;当两眼注视培墙上一个小黑点时,由于有眼外肌的调节,此点就都正好成像在两侧眼的黄斑上,于是在视觉中只“看到”一个点;此时如用手轻推一侧眼球的外侧,使此眼视轴稍作偏移,则这时此眼视网膜上的黑点像就要从黄斑部移开,落在与对侧视网膜像非相称的点上,于是会感到墙上有两个黑点存在,这就是复视现象。显然,在黄斑部以外,一眼的颞侧视网膜和另一眼的鼻侧视网膜互相对称;而一眼的鼻侧视网膜也与另正好的颞侧视网膜互相对称。
第三节 听觉器官
听觉的外周感受器官是耳,耳的适宜刺激是一定频率范围内的声波振动。耳由外耳、中耳和内耳迷路中的耳蜗部分组成。由声源振动引起空气产生疏密波,后者通过外耳道、鼓膜和听骨链的传递,引起耳蜗中淋巴液和基底膜的振动,使耳蜗科蒂器官中的毛细胞产生兴奋。科蒂器官和其中所含的毛细胞,是真正的声音感受装置,外耳和中耳等结构只是辅助振动波到达耳蜗的传音装置。听神经纤维就分布在毛细胞下方的基底膜中;振动波的机械能在这里转变为听神经纤维上的神经冲动。并以神经冲动的不同频率和组合形式对声音信息进入编码,传送到大脑皮层听觉中构,产生听觉。听觉对动物适应环境和人类认识自然有重要的意义;在人类,有声语言是互
通信息交流思想的重要工具。
因此,在耳的生理功能研究中主要解决的问题是:声音怎样通过外耳、中耳等传音装置传到耳蜗,以及耳蜗的感音装置如何把耳蜗淋巴液和基底膜的振动转变成为神经冲动。
一、人耳的听阈和听域
耳的适宜刺激是空气振动的疏密波,但振动的频率必须在一定的范围内,并且达到一定强度,才能被耳蜗所感受,引起听觉。通常人耳能感受的振动频率在16-20000Hz之间,而且对于其中每一种频率,都有一个刚好能引起听觉的最小振动强度,称为听阈。当振动强度在吸阈以上继续增加时,听觉的感受也相应增强,但当振动强度增加到某一限度时,它引起的将不单是听觉,同时还会引起鼓膜的疼痛感觉,这个限度称为最大可听阈。由于对每一个振动频率都有自己的听阈和最大或听阈,因而就能绘制出表示人耳对振动频率和强度的感受范围的坐标图,如图9-14所示。其中下方曲线表示不同频率振动的听阈,上方曲线表示它们的最大听阈,两得所包含的面积则称为听域。凡是人所能感受的声音,它的频率和强度的坐标都应在听域的范围之内。由听域图可看出,人耳最敏感的频率在1000-3000Hz之间;而日常语言的频率较此略低,语音的强度则在听阈和最大可听阈之间的中等强度处。
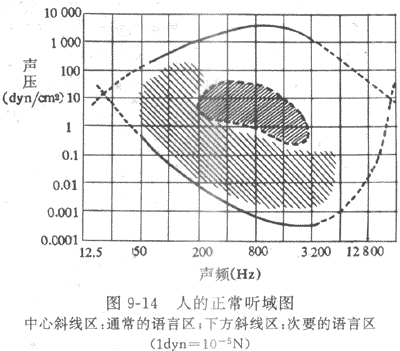
二、外耳和中耳的传音作用
(一)耳廓和外耳道的集音作用和共鸣腔作用
外耳由耳廓和外耳道组成。人耳耳廓的运动能力已经退化,但前方和侧方来的声音可直接进入外耳道,且耳廓的形状有利于声波能量的聚集,引起较强的鼓膜振动;同样的声音如来自耳廓后方,则可被耳廓遮挡,音感较弱。因此,稍稍转动头的位置,根据这时两耳声音强弱的轻微变化,可以判断音源的位置。
外耳首是声波传导的通路,一端开口,一端终止于鼓膜。根据物理学原理,充气的管道可与波长4倍管长的声波产生最大的共振作用;外耳道长约2.5cm,据此计算,它作为一个共鸣腔的最佳共振频率约在3500Hz附近;这样的声音由外耳道传到鼓膜时,其强度可以增强10倍。
(二)鼓膜和中耳听骨链增压效应
中耳包括鼓膜、鼓室、听骨链、中耳小肌和咽鼓管等主要结构,其中鼓膜、听骨链和内耳卵圆窗之间的关系如图9-15所示,它们构成了声音由外耳传向耳蜗的最有效通路。声波在到达鼓膜交,由空气为振动介质;由鼓膜经听骨链到达卵圆窗膜时,振动介质变为固相的生物组织。由于不同介质的声阻拦不同,理论上当振动在这些介质之间传递时,能量衰减极大,估计可达99%或更多。但由于由鼓膜到卵圆窗膜之间的传递系统的特殊力学特性,振动经中耳传递时发生了增压效应,补偿了由声阻挡不同造成的能量耗损。
鼓膜呈椭圆形,面积约50-90mm2,厚度约0.1mm。它不是一个平面膜,呈顶点朝向中耳的漏斗形。其内侧连锤骨柄,后者位于鼓膜的纤维层和粘膜层之间,自前上方向下,终止于鼓膜中心处。鼓膜很像电话机受话器中的振膜,是一个压力承受装置,具有较好的频率响应和较小的失真度,而且它的形状有利于把振动传递给位于漏斗尖顶处的锤骨柄。据观察,当频率在2400Hz以下的声波作用于鼓膜时,鼓膜都可以复制外加振动的频率,而且鼓膜的振动与声波振动同始同终,很少残余振动。

图9-15 人中耳和耳蜗关系模式图
点线表示鼓膜向内侧振动时各有关结构的移动情况
听骨链由锤骨、砧骨及镫骨依次连接而成。锤骨柄附着于鼓膜,镫骨脚板和卵圆窗膜相接,砧骨居中,将锤骨和镫骨连接起来,使三块听小骨形成一个两壁之间呈固定角度的杠杆。锤骨柄为长臂,砧骨长突为短臂。该械杆系统的特点是支点刚好在整个听骨链的重心上,因而在能量传递过程中惰性最小,效率最高。鼓膜振动时,如锤骨柄内移,则砧骨的长突和镫骨亦和锤骨柄作同方向的内移,如图9-15中点线所示。
中耳增压泖应主要有以下两个因素:一是由于鼓膜面积和卵圆窗膜的面积大小有差别,鼓膜振动时,实际发生振动的面积约55mm2,而卵圆窗膜的面积只有3.2mm2,如果听骨链传递时总压力不变,则作用于卵圆窗膜上的压强将增大55÷3.2=17倍;二是听骨链中杠杆长臂和短臂之比约为1.3:1,即锤骨柄较长,于是短臂一侧的压力将增大为原来的1.3倍。这样算来,整个中耳传递过程的增压效应为17×1.3=22倍。
与中耳传音功能有关的,还有中耳内的两条小肌肉,其中鼓膜张肌收缩时,可使锤骨柄和鼓膜内向牵引,增加鼓膜紧张度;镫骨肌收缩时,使镫骨脚板向外后方移动。强烈的声响气流经过外耳道,以及角膜和鼻粘膜受到机械刺激时,都可以反射性地引起这两块小肌肉的收缩,其结果是使鼓膜紧张,使各听小骨之间的边境更为紧张,导致吸骨链传递振动的幅度减小;阻力加大,总的效果是使中耳的传音效能有所减弱。据认为,这一反应可以阻止较强的振动传到耳蜗,对感音装置起到某种保护作用;但由于声音引起中耳肌的反射性收缩需经过十几个毫秒的潜伏期,故它们对突然发生的短暂爆炸声的保护作用不大。
(三)咽鼓管的功能
咽鼓管亦称耳咽管,它连通鼓室和鼻咽部,这就使鼓室内空气和大气相通,因而通过咽鼓管,可以平衡鼓室内空气和大气压之间有可能出现的压力差,这对于维持鼓膜的正常位置、形状和振动性能有重要意义。咽鼓管阻塞时,鼓室气体将被吸收,使鼓室内压力下降,引起鼓膜内陷。暂时的鼓膜内外压力差,常发生在外耳道内压力首先发生改变而鼓室内压力仍处于原初的状态,如飞机的突然升降长潜水等,此时如果不能通过咽鼓管使鼓室内压力外耳道压力(或大气压)取得平衡,就会在鼓膜两侧出现巨大的压力差。据观察,这个压力差如达到9.33-10.76kPa(70-80mmHg),将会引起鼓膜强烈痛疼;压力差超过24kPa(180mmHg)时,可能造成鼓膜破裂。咽鼓管在正常情况下其鼻咽部开口常处于闭合状态,在吞咽、打呵欠或喷嚏时由于腭帆张肌等肌肉的收缩,可使管口暂时开放,有利于气压平衡。
声音的骨传导 正常时听觉的引起,是由于声波经外耳道引起鼓膜的振动,再经听骨链和卵圆窗膜进入耳蜗,这一条声音传递地途径,称为气传导。此外,声波还可以直接引起颅骨的振动,再引起位于颞骨骨质中的耳蜗内淋巴的振动,这称为骨传导。骨传导正常时较气传导不敏感得多,几乎不能感到它的存在;能察知骨传导存在的一种方面是,把一个振动阒的音叉的柄直接和颇骨接触,这时人会感到一个稍有异样的声音;当这个声音减弱到听不到以后,再把音叉迅速移到耳廓前方,这时又能听到声音的存在。这个简单实验说明骨传导的存在,也说明正常时气传导较骨传导为灵敏。可以认为,骨传导在正常听觉的引起中作用微乎其微。不过临床上常通过检查患者气传导和骨传导受损的情况,判断听觉异常的产生部位和原因。
三、耳蜗的感音换能作用
耳蜗的作用是把传到耳蜗的机械振动转变成听神经纤维的神经冲动。在这一转变过程中,耳蜗基底膜的振动是一个关键因素。它的振动使位于它上面的毛细胞受到刺激,引起耳蜗内发生各种过渡性的电变化,最后引起位于毛细胞底部的传入神经纤维产生动作电位。
(一)耳蜗的结构要点
耳蜗是一条骨质的管道围绕一个骨轴盘旋21/2-23/4周而成。在耳蜗管的横断面上可见到两个分界膜,一为斜行的前庭膜,一为横行的基底膜,此两膜将管道分为三个腔,分别称为前庭阶、鼓阶和蜗管(图9-16)。前庭附在耳蜗底部与卵圆窗膜相接,内充外淋巴;鼓阶在耳蜗底部与圆窗膜相接,也充满外淋巴,后者在耳蜗顶部和前庭阶中的外淋巴相交通;蜗管是一个盲管,其中内淋巴浸浴着位于基底膜上的螺旋器的表面。螺旋器的构造极为复杂;在蜗管的横断面上的靠蜗轴一侧,可看到有一行内毛细胞纵向排列;在蜗管的靠外一侧,有3-5行外毛细胞纵向排列(参看图9-18);此外还有其他的支持细胞和存在于这些细胞间的较大的间隙,包括内、外隧道和Nuel间隙。需要指出的是,这些间隙中的液体在成分上和外淋巴一致,它们和蜗管中的内淋巴不相交通,但可通过基底膜上的小孔与鼓阶中的外淋巴相交通。这样的结构使得毛细胞的顶部与蜗管中的内淋巴相接触,而毛细胞的周围和底部则和外淋巴相接触。每一个毛细胞的项部表面,都有上百条排列整齐的听毛,其中较长的一些埋植在盖膜的冰胶状物质中,有些则只和盖膜接触。盖膜在内侧连耳蜗轴,外侧游离在内淋巴中。
(二)基底膜的振动和行波理论
当声流振动通过听骨链到达卵圆窗膜时,压力变化立即传给隔离蜗内液体和膜性结构;如果卵圆窗膜内移,前庭膜和基底膜也将下移,最后是鼓阶的外淋巴压迫圆窗膜外移;相反,当卵圆窗膜外移时,整个耳蜗内结构又作反方向的移动,于是形成振动。可以看出,在正常气传导的过程中,圆窗膜实际起着缓冲耳蜗内压力变化的作用,是耳蜗内结构发生振动的必要条件。有人用直接观察的方法,详细记录了声音刺激引起的基底膜振动的情况,这对于了解基底膜振动的形式,以及这种振动在耳蜗接受不同频率的声音刺激时有何差异,提供了可靠的依据。观察表明,基底膜的振动是以行波(traveling wave)的方式进行的,即内淋巴的振动首先是靠近卵圆窗处引起基底膜的振动,此波动再以行波的形式沿基底膜向耳蜗的顶部方向传播,就像人在抖动一条绸带时,有行波沿绸带向远端传播一样。下一步还证明,不同频率的声音引起的行波都从基底膜的底部,即靠近卵圆窗膜处开始,但频率不同时,行波传播的远近和最大行波的出现部位有气温同,如图9-17所示,;这就是振动频率愈低,行波传播愈远,最大行波振幅出现的部位愈靠近基底膜顶部,而且在行波最大振幅出现后,行波很快消失,不再传播;相反地,高频率声音引起的基底膜振动,只局限于卵圆窗附近。
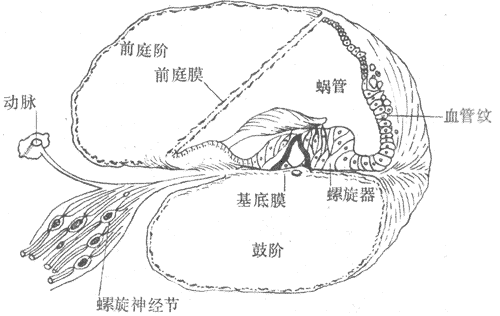
图9-16 耳蜗管的横断面图
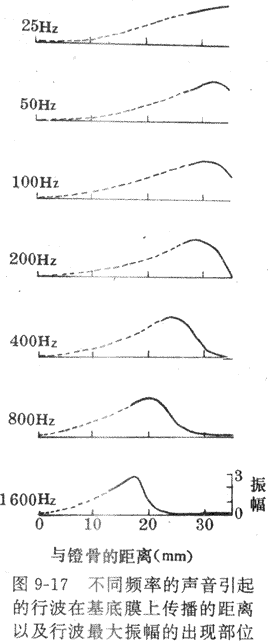
不同频率的振动引起的基底膜不同形式的行波传播,主要由基底膜的某些物理性质决定的。基底膜的长度在人约为30mm,较耳蜗略短,但宽度在靠近卵圆窗处只有0.04mm,以且逐渐加宽;与此相对应,基底膜上的蚴旋器的高度和重量,也随着基底膜的加宽而变大。这些因素决定了基底膜愈靠近底部,共振频率愈高,愈靠近顶部,共振频率愈低;这就使得低频振动引起的行波在向顶部传播时阻力较小,而高频振动引起的行波只限局在底部附近。
不同频率的声音引起的不同形式的基底膜的振动,被认为是耳蜗能区分不同声音频率的基础。破坏动物不同部位基底膜的实验和临床上不同性质耳聋原因的研究,都证明了这一结论,亦即耳蜗底部受时主要影响高频听力,耳蜗顶部受损时主要影响低频听力。不能理解,既然每一种振动频率在基底膜上都有一个特定的行波传播范围和最大振幅区,与这些区域有关的毛细胞和听神经纤维就会受到最大的刺激,这样,来自基底膜不同区域的听神经纤维的神经冲动及其组合形式,传到听觉中枢的不同部位,就可能引起不同音调的感觉。
基底膜的振动怎样使毛细胞受到刺激,如图9-18所示。毛细胞顶端的听毛有些埋在盖膜的胶状物中,有些是和盖膜的下面接触;因盖膜和基底膜的振动轴不一致,于是两膜之间有一个横向的交错移动,使听毛受到一个切向力的作用而弯曲(图9-18,下)。据研究,毛细胞听纤毛的弯曲,是耳蜗中由机械能转为电变化的第一步。
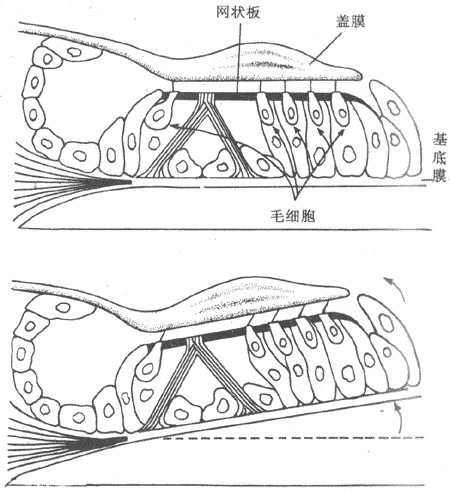
图9-18基底膜和盖膜振动时毛细胞顶部听毛受力情况
上:静止时的情况 下:基底膜在振动中上移时,因与盖膜之间的切向运动,听毛弯向蜗管外侧
(三)耳蜗的生物现象
在耳蜗结构中除了能记录到与听神经纤维兴奋有关的动作电位,还能记录到一些其他形式的电变化。在耳蜗未受到刺激时,如果把一个电极放在鼓阶外淋巴中,并接地使之保持在零电位,那么用另一个测量电极可测出蜗管内淋巴中的电位为+80mV左右,这称为内淋巴电位。如果将此测量电极刺入毛细胞膜内,则膜内电位为-70?/FONT>-80mV。毛细胞顶端膜外的浸浴液为内淋巴,则该处毛细胞内(相当于-80mV)和膜外(相当于+80mV)的电位差当为160mV;而在毛细胞周围的浸浴液为外淋巴(电位相当于零),该处膜内外的电位差只有80mV左右;这是毛细胞静息电位和一般细胞不同之处。据实验分析,内淋巴中正电位的产生和维持,同蜗管外侧壁处的血管纹结构的细胞活动有直接关系(图9-16),并且对缺 O2非常敏感;有人发现,血管纹细胞的膜含有大量活性很高的ATP酶,具有“钠泵”的作用,它们可依靠分解ATP获得能量,将血浆中的K+泵入内淋巴,将内淋巴中的Na+泵入血浆,但被转运的K+担超过了Na+的量,这就使内淋巴中有大量K+蓄积,因而使内淋巴保持了较高的正电位;缺O2使ATP的生成受阻,也使Na+泵的活动受阻,因而使内淋巴的正电位不能维持。
当耳蜗接受声音刺激时,在耳蜗及其附近结构又可记录到一种特殊的电波动,称为微音器电位。这是一种交流性质的电变化,在一定的刺激强度范围内,它的频率和幅度与声波振动完全一致(图9-19);这一现象正如向一个电话机的受话器或微音器(即麦克风)发声时,它们可将声音振动转变为波形类似的音频电信号一样,这正是把耳蜗的这种电变化称为微音器电位的原因。事实上,如果对着一个实验动物和耳廓讲话,同时在耳蜗引导它的微音器电位,并将此电位经放大后连接到一个扬声器,那么扬声器发出的声音正好是讲话的声音!这一实验生动地说明,耳蜗在这里起着类似微音器的作用,能把声波变成相应的音频电信号。微音器电位的其它一些特点是:潜伏期极短,小于0.1ms;没有不应期;对缺O2和深麻醉相对地不敏感,以及它在听神经纤维变性时仍能出现等。
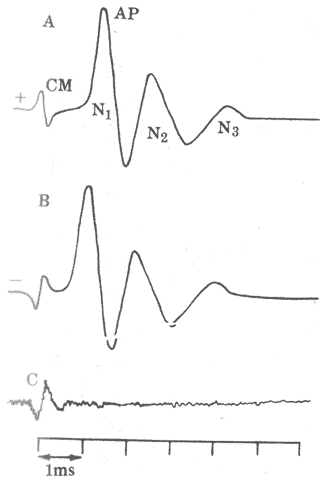
图9-19 由短声刺激引起的微音器电位和听神经动作电位
CM:微音器电位 AP:耳蜗神经动作电位(包括N1、N2、N3三个负电位)
A与B对比表明,声音位相改变时,微音器电位位相倒转,但神经动作
电位位相没有变化 C:在白噪音作用下,AP消失,CM仍存在
用微电极进入毛细胞的细胞内电变化记录的实验证明,所谓微音器电位就是多个毛细胞在接受声音刺激时产生的感受器电位的复合表现;在记录单一毛细胞跨膜电位的情况下,发现听毛只要有0.1。的角位移,就可引起毛细胞出现感受器电位,而且电位变化的方向与听毛受力的方向有关,亦即此电位既可是去极化的;这就说明了为什么微音器电位的波动同声波振动的频率和幅度相一致。
由于听毛的角位移和产生感受器电位之间只有一极短潜伏期,因而认为后者的产生是由于毛细胞顶部膜中有机械门控通道的存在,听毛受力引起该处膜的轻微变形,就足以改变这种通道蛋白质的功能状态,引起跨膜离子移动和相应的电位反应。在毛细胞,它的感受器电位可引起细胞基底部的递质(可能是谷氨酸和门冬氨酸)释放量的改变,进而引起分布在附近的耳蜗传入纤维产生动作电位,传向听觉高级枢,产生听觉。至于内毛细胞和外毛细胞在功能上有何不同,有人首先注意到它们所接受的传入纤维的数目有极大差异。据计算,人一侧耳蜗内毛细胞的总数约为3500个,外毛细胞则有约15000个,但来自螺旋神经节的约32000条听神经传入纤维中约有90%分布到内毛细胞的底部,这说明一个内毛细胞可接受多条传入纤维的分布,而多个外毛细胞才能接受一个传入纤维的轴突分支。因此一般认为,内毛细胞的作用是把不同频率的声音振动转变为大量分布在它们底部的传入纤维的神经冲动,向中枢传送听觉信息,而息细胞的作用近年来却发现有些特殊。有人发现毛细胞在基底膜振动和听毛受力而出现微音器电位时,此细胞可产生形体长短的快速改变,超极化引起细胞伸长,去极化引起细胞缩短,它们的形体改变因此也和外来声音振动的频率和振幅同步。据认为,外毛细胞的这种形体改变可以使所在基底膜部分原有的振动增强,亦却对行经该处的行波起放大作用,这显然使位于该部分基底膜上的内毛细胞更易受到刺激,提高了对该振动频率的敏感性。外毛细胞因膜内外电位差改变引起的机制尚不清楚,但这使得基底膜不仅仅是以固定的结构“被动”地对外界的振动产生行波,它还可以“主动”地增强行波的振动幅度。
四、听神经动作电位
听神经纤维的动作电位,是耳蜗对声音刺激一一系列反应中最后出现的电变化,是耳蜗对声音刺激进行换能和编码作用的总结果,中枢的听觉感受只能根据这些传入来引起。图9-19中的N1、N2、N3……是从整个吸神经上记录到复合动作电位,并非由单一听神经纤维的兴奋所产生。用不同频率的纯音刺激耳蝇,同时检查不同的单一听神经纤维的冲动发放情况,就可检查行波理论是否正确,也可阐明耳蜗编码作用的某些特点。仔细分析每一条听神经纤维的放电牧场生和声音频率之间的关系时,发出如果声音强度够大时,同一纤维常可对一组频率相近的纯音刺激起反应,但如果将声音强度逐渐减弱,则可找到一个纤维的最佳反应频率,即当别的刺激频率都因强度太弱而不引起动作电位时,该频率仍能引起。每一条纤维的最佳反应频率的高低,决定于该纤维末稍的基底膜上分布位置,而这一部分正好是该频率的声音所引起的最大振幅行波的所在位置。由这类实验得出的初步结论是,当某一频率的声音强度较弱时,神经信号由少数对该频率最敏感的神经纤维向中枢传递,当这一频率的声音强度增大时,除了能引起上述纤维兴奋外,还能引起更多一些其最佳反应频率与该频率相近的神经纤维也发生兴奋,使更多的纤维参加到传音的频率及其强度的行列里来,结果是由这些纤维传递的神经冲动,共同向中枢传递这一声音的频率及其强度的信息。在自然情况下,作用于人耳的声音捱牟频率和强度的变化是十分复杂的,因此基底膜的振动形式和由此而引起听神经纤维的兴奋及其组合也是十分复杂的;人耳可以区别不同音色,其基础可能亦在于此。
第四节 前庭器官
内耳迷路中除耳蜗外,还有三个半规管、椭圆囊和球囊,后三者合称为前庭器官,是人体对自身运动状态和头在空间位置的感受器。当机体进行旋转或直线变速运动时,速度的变化(包括正、负加速度)会刺激三个半规圆或椭圆囊中的感受细胞;当头的位置和地球引力的作用方向出现相对关系的改变时,就会刺激球囊中的感受细胞。这些刺激引起的神经冲动沿第八脑神经的前庭支传向中枢,引起相应的感受和其他效应。
一、前庭器官的感受装置和适宜刺激
前庭器官的感受细胞都称为毛细胞,具有类似的结构和功能。这些毛细胞通常在顶部有60-100条纤细的毛,按一定的形式排列;其中有一条最长,位于细胞顶端的一侧边缘处,称为动毛,其余的毛较短,占据了细胞顶端的大部分区域,称静毛。图9-20是在一个半规管壶腹中的毛细胞上所做的实验,当动毛和静毛都处于自然状态时,细胞膜内外存在着约-80mV的静息电位,同时在与此毛细胞相接触的神经纤维上有中等频率的持续放电;此时如果用外力使毛细胞顶部的纤毛由静毛所在一侧倒向动毛一侧,可看到细胞的静息电位去极化到约-60mV的水平,同时有神经纤维冲动发放频率的增加;与此相反,当外力使纤毛弯曲的方向由动毛一侧向静毛一侧时,可看到细胞静息电位向超极化的方向转变,而神经纤维上的冲动发放频率也变得比纤毛处自然不受力状态时为小.这是迷路器官中所有毛感受外界刺激时的一般规律,其换能机制与前面讲到的耳蜗毛细胞类似。在正常情况下,由于各前庭器官中毛细胞的所在位置和附属结构的不同,便得不同形式的变速运动都能以特定的方式改变毛细胞纤毛的倒向,使相应的神经纤维的冲动发放频率发生改变,把机体运动状态和头在空间位置的信息传送到中枢,引起特殊的运动觉和位置觉,并出现各种躯体和内脏功能的反射性改变。
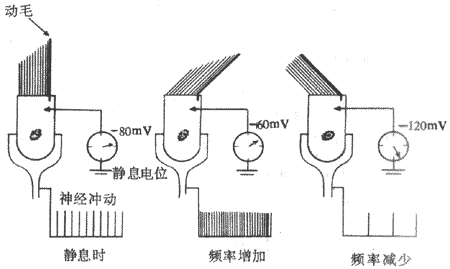
图9-20 前庭器官中毛细胞顶部纤毛受力情况影响 细胞静息电位和神经纤维冲动发放频率的实验示意图
三个半规管的形状大致相同,但各处于一个平面上,这三个平面又互相垂直。每个半规管约占2/3个圆周,一端有一个相对膨大的壶腹。两侧的水平半规管同时在一个平面上,如果人在直立时头前倾30℃,则此平面正好与地面平行;当两臂平举而肘关节呈半屈状态时,此时手臂的方位即相当于水平半规管的方位,两个拳头的位置就相当于两侧壶腹的位置。壶腹内有壶嵴,它的位置和半规管的轴垂直;在壶嵴中有一排毛细胞,面对管腔,而毛细胞顶部的纤毛又都埋植在一种胶质性的圆顶形终帽之中。毛细胞上动毛和静毛的相对位置是固定的,例如在水平半规管内,当充满管腔的内淋巴由管腔向壶腹的方向移动时,正好能使壶嵴中毛细胞顶部的静毛向动毛一侧弯曲,于是引起该侧壶腹的传入神经向中枢发放大量的神经冲动。
水平半规管的结构特点,使它能感受人体以身体长轴为轴所作的旋转变速运动。旋转开始时,由于管腔中内淋巴的惯性作用,它的起动将晚于人体和管本身的运动,因此当人体向左旋转时,左侧水平半规管中的内淋巴将压向壶腹的方向,使该侧毛细胞兴奋而产生较多的神经冲动;与此同时,右侧水平半规管中的内淋巴压力作用方向正好是离开壶腹,于是由该侧壶腹传向中枢的冲动减少。人脑正是根据来自两侧水平半规管传入信号的不同,“判定”人体是否开始旋转和向何方旋转的。当旋转变为匀速旋转时,管腔中内淋巴与整个管同步运动,于是两侧壶腹中的毛细胞都处于不受力状态,中枢获得的信息与不进行旋转时无异。但当人体停止旋转时,内淋巴运动的停止又由于惯性作用晚于管本身,于是两侧壶腹中的毛细胞又有受力情况的改变,其受力方向和冲动发放情况正好与旋转开始时相反。内耳迷路中尚有其他两对半规管,可以接受和它们所处平面方向一致的旋转变速运动的刺激。
在椭圆囊和球囊,毛细胞存在于囊斑结构中,其纤毛则埋植在一种称为耳石膜的结构内。耳石膜是一块胶质板,内含耳石,主要由蛋白质和碳酸钙所组成,比重大于内淋巴,因而也有较大的惯性。椭圆囊和球囊的不同,在于其中囊斑所在的平面和人体的相对关系不一样。人体在直立位时,椭圆囊中囊斑所处平面呈水平,囊斑表面分布的毛细胞顶部朝上,耳石膜在纤毛上方;球囊与此不同,其中囊斑所处平面在人体直立时位置和地面垂直,毛细胞和第十毛由囊斑表面向水平方向伸出,耳石膜悬在纤毛外侧,与囊斑相平行。仔细检查两个囊斑平面上分布着的各毛细胞顶部静毛和动毛的相对位置关系时,发现这在每一个毛细胞几乎都不相同(图9-21)。毛细胞纤毛的这种配置,使得它们有可能分辨人体在囊斑平面所作的各种方向的直线变速运动。例如,当人体在水平方向以任何角度作直线变速运动时,由于耳石膜的惯性,在椭圆囊囊斑上总会有一些毛细胞由于它们的静毛和动毛的独特的方位,正好能发生静毛向动毛侧的最大弯曲,于是由此引起的某些特定的传入神经纤维的冲动发放增加,引起机体产生进行着某种方向的直线变速运动的感觉。球囊囊斑上的毛细胞,则由于类似的机制,可以感受头在空间位置和重力作用方向之间的差异,因而可以“判断”头以重力作用方向为参考点的相对位置变化。
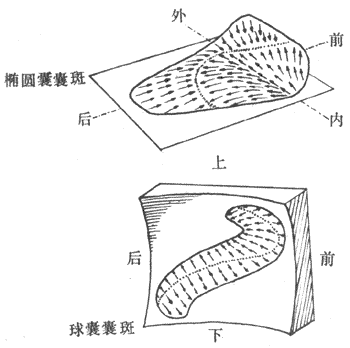
图9-21 椭圆囊和球囊中囊斑的位置以及毛细胞顶部纤毛的排列方向
箭头所指方向是该处毛细胞顶部动毛所在位置,箭尾是同一细胞的静毛所在位置,
当机体所作直线加速运动的方向与某一箭头的方向一致时,该箭头所代表的毛细胞
表面静毛向动毛侧的弯曲最明显,与此毛细胞有关的神经纤维有最大频率的冲动发放
二、前庭反应和眼震颤
来自前庭器官的传入冲动,除与运动党内和位置觉的引起有关外,还引起各种姿势调节的反射和植物性功能的改变。延髓动物(见第十章)表面的某些状态反射和中礅动物的翻正反射,就与前庭器官的传入冲动有关。人体在前庭器官受到刺激时,也会出现一些躯体调节反应,如人乘车而车突然加速时,会有背肌紧张增强而后仰,车突然减速时又有相反的情况;当电梯突然上升时,肢体伸肌抑制而屈曲,下降时伸肌紧张加强而伸直,等等。
前庭反应是中最特殊的是躯体旋转运动时出现的眼球的特殊运动,称为眼震颤,常被用来判断前庭功能是否正常。眼震颤主要由半规管的刺激引起,而且眼震颤的方向也由于受刺激半规管的不同而不同。当人体头部前倾30℃而围绕人体垂直轴旋转时,主要是两侧的水平半规管壶嵴毛细胞有刺激强度的改变,这时出现的也是水平方向的眼震颤。具体情况是,当旋转开始时,如果是向左侧旋转,则是左侧壶嵴的毛细胞受刺激增强而右侧正好相反,这时出现两侧眼球缓慢向右侧移动,这称为眼震颤的慢动相;当慢动相使眼球移动到两眼裂右侧端而不能再移时,又突然返回到眼裂正中,这称为眼震颤的慢动相;此后再出现新的慢动相和快动相,返复不已,这就是眼震颤。当旋转变为匀速运动时,旋转虽在继续,但由于两则壶嵴所受压力一样,于是眼球不再震颤而居于眼裂正中。只有当旋转停止而出现减速时,内淋巴又由于惯性作用而不能立刻停止运动,于是两侧壶嵴又再现所受压力的不同,但情况正好与旋转开始时相反,于是又引起一阵由方向相反的慢动相和快动相组成的眼震颤(图9-22)。临床和特殊从业人员常进行眼震颤试验以判断前庭功能是否正常。在同样条件下震颤时间过长或过短,说明前庭功能有过敏或减弱,前庭器官受到过强或过长刺激,或刺激未过量而前庭功能过敏时,常会引起恶心、呕吐、眩晕、皮肤苍白等现象,称为前庭植物神经性反应,具体表现为晕船、晕车和航空病。
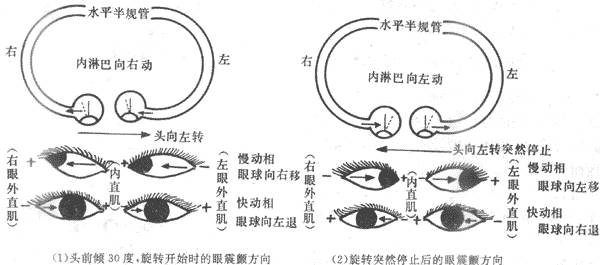
(1)头前倾30℃,旋转开始时的眼震颤方向(2)旋转突然停止后的眼震颤方向
图9-22 旋转变速运动时两侧水平半规管壶嵴毛细胞受刺激情况和眼震颤方向示意图
第五节 嗅觉和味觉
一、嗅觉感受器和嗅觉的特点
嗅觉感受器位于上鼻道及鼻中隔后上部的嗅上皮,两侧总面积约5cm2。由于它们的位置较高平静呼吸时气流不易到达。因此在嗅一些不太显着的气味时,要用力吸气,使气流上冲,才能到达嗅上皮。嗅上皮含有三种细胞,即主细胞、支持细胞和基底细胞。主细胞也称呼嗅细胞(图9-23),呈圆瓶状,细胞顶端有5-6条短的纤毛,细胞的底端有长突,它们组成嗅丝,穿过筛骨直接进入嗅球。嗅细胞的纤毛受到存在于空气中的物质分子刺激时,有神经冲动传向嗅球,进而传向更高级的嗅觉中枢,引起嗅觉。
不同动物的嗅觉敏感程度差异很大,同一动物对不同有气味物质的敏感程度也不同。嗅上皮和有关中枢究竟怎样感受并能区分出多种气味,目前已有初步了解。有人分析了600种有气味物质和它们的化学结构,提出至少存在7种基本气味;其他众多的气味则可能由这些基本扬眉吐气的组合所引起。这7种基本气味是:樟脑味、麝香味、花卉味、薄荷味、乙醚味、辛辣味和腐腥味;他们发现,大多数具有同样气味的物质,具有共同的分子结构有特殊结合能力的受体蛋白(理论上至少有7种),这种结合可通过G-蛋白而引起第二信使类物质的产生,最后导致膜上某种离子通道开放,引起Na+、K+等离子的跨膜移动,在嗅细胞的胞体膜上产生去极化型的感受器电位,后者在轴突膜上引起不同频率的动作电位发放,传入中枢。用细胞内记录法检查单一嗅细胞电反应的实验发现,每一个嗅细胞只对一种或两种特殊的气味坡反应;还证明嗅球中不同部位的细胞只对某种特殊的气味起反应。嗅觉系统也其他感觉系统类似,不同性质的气味刺激有其相对专用的感受位点和传输线路;非基本气味则由于它们在不同线路上引起的不同数量冲动的组合特点,在中枢引起特有的主观嗅觉感受。
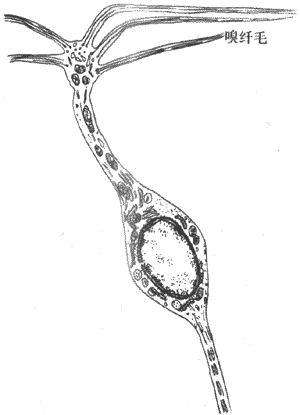
图9-23 嗅细胞(双极细胞)
二、味觉感受器和味觉的特点
味觉的感受器是味蕾,主要分布在舌背部表面和知缘,口腔和咽部粘膜的表面也有散在的味蕾存在。儿童味蕾较成人为多,老年时因萎缩而逐渐减少。每一味蕾由味觉细胞和支持细胞组成(图9-24)。味觉细胞顶端有纤毛,称为味毛,由味蕾表面的孔伸出,是味觉感受的关键部位。
舌表面不同部分对不同味刺激的敏感程度不一样。在人,一般是舌尖部对甜味道比较敏感,舌两侧对酸味比较敏感。舌两侧前部对咸味比较敏感,而软腭和舌根部对苦味比较敏感。味觉的敏感度往往受食物或刺激物本身温度的影响。在20-30℃之间,味觉的敏感度最高。另外,味觉的辨别能力也受血液化学成分的影响,例如,动物实验中正常大鼠能辩出1:2000的氯化钠深夜,而切除上腺皮质的大鼠,可能是由于血液中低Na+,可辨别出1:33000的氯化钠深夜,主动选饮这种含盐多的深夜。因此,味觉的功能不仅在于辨别不同的味道,而且与营养物的摄取和内环境恒定的调节也有关系。
人和动物 味觉系统可以感受和区分出多种味道;但很早以前就知道,众多的味道是由四种基本的味觉组合而成的,这就是甜、咸、酸和苦。不同物质的味道与它们的分子结构的形式有关,便也有例外。通常NaCI能引起典型的咸味;甜味的引起与葡萄糖的主体结构有关;而奎宁和一些有毒植物的生物碱的结构能引起典型的苦味。有趣的是,这4种基本味觉的换能或跨膜信号的转换机制并不一样,如咸和酸的刺激要通过特殊化学门控通道,甜味的引起要通过受体、G-蛋白和第二信使系统,而苦味则由于物质结构不同而通过上述两种形式换能。和前面讲过的嗅觉刺激的编码过程类似,中枢可能通过来自传导四种基本味觉的专用神经通路上的神经信号和不同组合来“认知”这些基本味觉的以外的多种味觉。
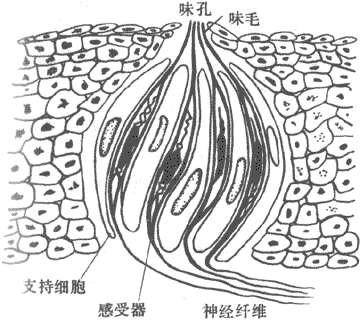
图9-24 味蕾的结构
第六节 皮肤感受
皮肤内分布着多种感受器,能产生多种感觉。一般认为皮肤感觉主要有四种,即触觉、冷觉、温觉和痛觉。用不同性质的点状刺激仔细检查人的皮肤感觉时发现,不同感觉的感受区在皮肤表面呈互相独立的点状分布;如用纤细的毛轻触皮肤表面时,只有当某些特殊的点被触及时,才能引起触觉。用类似的方法,可找到冷觉点、热点和痛点等。用组织学方法曾发出皮肤中有大量游离的感觉神经末稍和种种特殊形式的感觉小体(图9-25),因此曾推测每一种性质的感觉应当同某一特定形式的感受结构相对应;但近年来的实验结果不支持这样的观点,例如,同样的游离神经末稍可以在不同的感觉点下方发现,而触点下方可以找到游离神经末稍或种种特殊形式的感觉小体。看来,不同感受器的功能特异性应到膜的分子水平上支找,各种附属结构可能只影响感受末梢敏感性的高低和适应性的快慢。皮肤感受器的换能机制除触-压觉是通过机构门控通道外,其他尚不清楚。
触觉是微弱的机械刺激兴奋了皮肤浅层的触觉感受器引起的,压觉是指较强的机械刺激导致深部组织变形时引起的感觉,两者在性质上类似,可统称为触-压觉。触点在皮肤表面会布密度和该部位对触觉的敏感程度成正比,如颜面、口唇、指尖等处密度较高,手背、背部密度较低。皮肤在接受每秒5-40次的机械振动刺激时,还可引起振动觉,据认为这与触觉感受器有关。与触觉有关的传入纤维既有髓的Ⅱ、Ⅲ类纤维,也有纤细的N类无髓纤维。
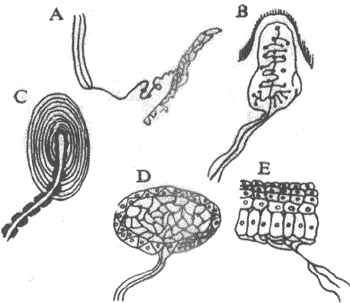
图9-25 皮肤中的各种感受装置
A:Ruffini小体 B:Meissner小体 C:环层小体即Pacini小体
D:Krause球 E:皮肤的游离神经末梢
冷觉和温觉合称温度觉,这起源于两种感受范围不同的温度感受器,因为冷不能构成一种能量长工。冷感受器在皮肤温度低于30℃时开始引起冲动发放,热感受器在超过30℃时开始引起冲动发放,47℃时频率最高。一般皮肤表面冷点约较热点多4-10倍;冷点下方主要分布有游离神经末梢,由Ⅲ类纤维传导传入冲动;热感受器可能也主要是游离神经末梢,传导纤维以N类为主。
痛觉是由有可能损伤或已经造成皮肤损伤的各种性质的刺激所引起的,它们除引起不愉快的痛苦感觉外,尚伴有强烈的情绪反应。
参考资料
1.杨雄里.光感受机制研究的重要突破.生理科学进展1987;18:1-14
2.Dowling JE,吴淼鑫,杨雄里译。视网膜。上海:上海医科大学出版社,1989
3.缪天荣,对数视力表及5分记录法。中华眼科杂志1966;13:96-104
4.Berne RM,Levy ML.Physiology 3rded,CV Mosby Co,St Louis,1993
5.Roper SD.The cell biology ofvertebrate.Ann Rev Neurosci 1989;12:329-353
6.Daw NW,Brunken WJ,ParkinsnD.The funcfion of synaptic transmitters in the retina.Ann Rev Neurosci1989;12:205-225
7.Dallos P.The active cochlea.JNeurosci 1992;12:4575-4585
8.Kinnamon SC,CummingsTA.Chemosensory transduction mechanisms in taste.Ann Rev Physiol1992;54:715-731
9.Von Beseky G.Experiments inHearing.McGraw-Hill Publishing Co,New York,1960
10.Zufall F,Firestein S,Shepherd G.Cyclicnucleotide-gated ion channels and sensory transduction in olfactory receptorneurons.Ann Rev Biophys and Biomol Structure 1944;23:577-607

 中医世家
中医世家 浦 标 网
浦 标 网 河南大学精品课程
河南大学精品课程 图书资料室
图书资料室