第八章索引
肾是维持机体内环境相对稳定的最重要的器官之一。通过尿的生成和排出,①排除机体的大部分代谢终产物以及进入体内的异物;②调节细胞外液量和渗透压;③保留体液中的重要电解质如钠、钾、碳酸氢盐以及氯离子等,排出氢离子,维持酸碱平衡。
尿的生成包括肾小球的滤过,肾小管和集合管的重吸收以及它们的分泌三个基本过程。
本章主要阐述肾的尿生成过程及其调节机制,以及输尿管和膀胱的排尿活动。
第一节 肾的功能解剖和肾血流量
一、肾的功能解剖
(一)肾单位和集合管
肾单位是肾的基本功能单位,它与集合管共同完成泌尿功能。人的两侧肾约有170-240万个肾单位,每个肾单位包括肾小体和肾小管部分(图8-1)。肾小体包括肾小球和肾小囊两部分。肾小球是一团毛细血管网,其峡谷端分别与入球小动脉和出球小动脉相连(图8-2)。肾小球的包囊称为肾小囊。它有两层上皮细胞,内层(脏层)紧贴在毛细血管壁上,外层(壁层)与肾小管壁相连;两层上皮之间的腔隙称为囊腔,与肾小管管腔相通。血浆中某些成分通过肾小球毛细血管网向囊腔滤出;滤出时必须通过肾小球毛细血管内皮细胞、基膜和肾小囊脏层上皮细胞,这三者构成滤过膜(图8-3)。

图8-1 肾单位示意图

图8-2 肾小球、肾小囊穿刺和近球小体示意图方框示近球小体

图8-3 滤过膜示意图
肾小管由近球小管、髓袢和远球小管三部分组成。近球小管包括近由小管和髓袢降支粗段。髓袢由髓袢降支和髓袢升支组成;前者包括髓袢降支粗段(也是近球小管的组成部分)和降支细段;后者是指髓袢升支细段和升支粗段(也是远球小管的一部分)。远球小管包括髓袢升支粗段和远曲小管。远曲小管末端怀集合管相连。
集合管不包括在肾单位内,但在功能上和远球小管密切相关,它在尿生成过程中,特别是在尿液浓缩过程中起着重要作用,每一集合管接受多条远内小管运来的液体。许多集合管又汇入乳头管,最后形成的尿液经肾盏、肾盂、输尿管而进入膀胱,由膀胱排出体外。
(二)皮质肾单位和近髓肾单位
肾单位按其所在部位不同,可分为皮质肾单位和近髓肾单位(髓旁肾单位)两类(图8-4)。
皮质肾单位主要分布于外皮质层和中皮质层。人肾的皮质肾单位约占肾单位总数的85%-90%。这类肾单位的肾小球体积较小;入球小动脉的口径比出球小动脉的粗,两者口径之比约为2:1。出球小动脉进一步再分为毛细血管后,几乎全部分布于皮质部分的肾小管周围。这类肾单位的髓袢甚短,只达外髓质层,有的甚至不到髓质。

图8-4 肾单位和肾血管的示意图
处于肾皮质不同部位的肾单位和肾血管的结构显著不同
近髓肾单位分布于靠近髓质的内皮质层,在人肾约占肾单位中的10%-15%。这类肾单位的肾小球体积较大;其髓袢甚长,可深入到内髓质层,有的甚至到达乳头部。出球小动脉不仅形成缠绕邻近的近曲小管或远曲小管的网状毛细血管,而且还形成细而长的U字形直小血管。直小血管可深入到髓质,并形成毛细血管网包绕髓袢升支和集合管。近髓肾单位和直小血管的这些解剖特点,决定了它们在尿的浓缩与稀释过程中起着重要作用。
(三)近球小体
近球小体(juxtaglomerular apparatus)由颗粒细胞、系膜(间质)细胞和致密斑三者组成(图8-2)。颗粒细胞是位于入球小动脉的中膜内的肌上皮样细胞,内含分泌颗粒,分泌颗粒内含肾素。系膜细胞是指入球小动脉和出球小动脉之间的一群细胞,具有吞噬功能。致密斑位于远曲小管的起始部分,此处的上皮细胞变为主柱状细胞,局部呈现斑纹隆起,称为致密斑,致密斑与入球小动脉和出球小动脉相接触。致密斑可感受小管液中NaCI含量的变化,并将信息传递至颗粒细胞,调节肾素的释放。
近球小体主要分布在皮质肾单位,因而皮质肾单位含肾素较多,而近髓肾单位则几乎不含肾素。肾素分布的这种差异,也提示两种肾单位在功能上有所不同。
(四)肾的神经支配
肾交感神经主要从胸12至腰12脊髓发出,其纤维经腹腔神经丛支配肾动脉(尤其是入球小动脉和出球小动脉的不滑肌)、肾小管和释放肾素的颗粒细胞。肾交感神经末稍释放去甲肾上腺素,调节肾血流量、肾小球滤过率、肾小管的重吸收和肾素释放。有人认为,未发现肾有副并感神经支配,肾的各种感受器可经肾神经传入脊髓,并从脊髓投射到中枢的不同部位。
(五)肾的血液供应
肾动脉由腹主动脉垂直分出,其分支经叶间动脉→弓形动脉→小叶间动脉→入球小动脉。每支入球小动脉进入肾小体后,又分支成肾小球毛细血管网,后者汇集成出球小动脉而离开肾小体。出球小动脉再次分成毛细血管网,缠绕于肾小管和集合管的周围。所以,肾血液供应要经过两次毛细血管网,然后才汇合成静脉,由小叶间静脉→弓形静脉→叶间静脉→肾静脉(图8-4)。
肾小球毛细血管网介于入球小动脉和出球小动脉之间,而且皮质肾单位入球小动脉的口径比出球小动脉的粗1倍。因此,肾小球毛细血管内血压较高,有利于肾小球的滤过作用;肾小管周围的毛细血管网的血压较低,可促进肾小管的重吸收。
二、肾血液循环的特征
肾的血液供应很丰富。正常成人安静时每分钟有1200ml血液流过两侧肾,相当于心输出量的1/5-1/4左右。其中约94%的血液分布在肾皮质层,5%-6%分布在外髓,其余不到1%供应内髓,通常所说的肾备注量主要指肾皮质血流量。
肾血流量的调节包括肾血流量的自身调节和神经体液调节。
(一)肾血流量的自身调节
肾血流量的自身调节表现为动脉血压在一定范围内变动时,肾血流量仍然保持相对恒定(图8-5)。离体肾实验观察到,当肾动脉的灌注压(相当于体内的平均动脉压)由2.7kPa(20mmHg)提高到10.7kPa(80mmHg)的过程中,肾血流量将随肾灌注压的升高而成比例地增加;而当灌注压在10.7-247kPa(80-180mmHg)范围内变动时,肾血流量又将随灌注压的长高而增加。这种不依赖肾外神经支配使肾血流量在一定的血压变动范围内能保持不变的现象,表明它具有自身调节。一般认为,自身调节只涉及肾皮质的血流量。

图8-5 肾血流量和肾小球滤过率的自身调节
RPF:肾血浆流量 GFR:肾小球滤过率
关于自身调节的机制,有人提出肌源学说来解释。此学说认为,当肾灌注压增高时,血管平滑肌因灌注压增加而受到牵张刺激,这使得不滑肌的紧张性加强,血管口径相应地缩小,血流的阻力便相应地增大,保持肾血流量稳定;而当灌注压减小时则发生相反的变化。由于在灌注压低于10.77kPa(80mmHg)时,平滑肌已达到舒张的极限;而灌注压高于247kPa(180mmHg)时,平滑肌又达到收缩的极限。因此,在10.7kPa(80mmHg)以下和24kPa(180mmHg)以上时,肾血流量的自身调节便不能维持,肾血流量将随血压的变化而变化。只有在10.7-24kPa(80-180mmHg)的血压变化范围内,入球小动脉平滑肌才能发挥自身调节作用,保持肾血流量的相对恒定。如果用罂粟碱、水合氯醛或氰化钠等药物抑制血管平滑肌的活动,自身调节便告消失。此外,还不能完全排除其它因素如肾内组织液压力、代谢产物等因素在肾血流量自身调节中的作用。通过肾血流量自身调节,使肾小球滤守率不会因血压波动而改变,维持肾小球滤过率相对恒定(图8-5)。
(二)肾血流量的神经和体液调节
肾血流量的神经、体液调节使肾血流量与全身的血液循环调节相配合。肾交感神经活动加强时,引起肾血管收缩,肾血流量减少。影响肾交感神经活动的因素已在心血管反射中作了详述(见第四章)。
肾上腺素与去甲肾上腺素都能使肾血管收缩,肾血流量减少。血管升压素和血管紧张素等也能使肾血管收缩;前列腺素可使肾血管扩张。
总之,在通常情况下,在一般的血压变化范围内,肾主要依靠自身调节来保持血流量的相对稳定,以维持正常的泌尿功能。在紧急情况下,全身血液将重新分配,通过交感神经及肾上腺素的作用来减少肾血流量,使血液分配到脑、心脏等重要器官,这对维持脑和心脏的血液供应有重要意义。
第二节 肾小球的滤过功能
循环血液经过肾小球毛细血管时,血浆中的水和小分子溶质,包括少量分子量较小的血浆蛋白,可以滤入肾小囊的囊腔而形成滤过液。用微穿刺法实验证明,肾小球的滤过液就是血浆中的超滤液。
微穿刺法是利用显微操纵仪将外径6-10μm的微细玻璃插入肾小体的囊腔中。在与囊腔相接部位的近球小管内,注入石蜡油防止起滤液进入肾小管。用微细玻璃管直接抽到囊腔中的液体进行微量化学分析(图8-2)。分析表明,除了蛋白质含量甚少之外,各种晶体物质如葡萄糖、氯化物、无机磷酸盐、尿素、尿酸和肌酐等的浓度都与血浆中的非常接近,而且渗透压及酸碱度也与血浆的相似,由此证明囊内液确是血浆的超滤液。
单位时间内(每分钟)两肾生成的超滤液量称为肾小球滤过率(glomerularfiltration rate,GFR)。据测定,体表面积为1.73m2的个体,其肾小球滤过率为125ml/min左右。照此计算,两侧肾每一昼夜从肾小球滤出的血浆总量将高达180L。此值约为体重的3倍。肾小球滤过率和肾血浆流量的比例称为滤过分数(filtration fraction)。经测算,肾血浆流量为66ml/min,所以滤过分数为:125/660×100=19%。滤过分数表明,流经肾的血浆约有1/5幔有小球小茁到囊腔中。肾小球滤过率大小决定于滤过系数(Kf)(即滤过膜的面积及其通透性的状态)和有效滤过压。肾小球滤过率=Kf×PUF,PUF表示有效滤过压。
一、滤过膜及其通透性
人体两侧肾全部肾小球毛细血管总面积估计在1.5m2以上,这样大的滤过面积有利于血浆的滤过。在正常情况下,人两肾的全部肾小球滤过面积可以保持稳定。但是在急性肾小球肾炎时,由于肾小球毛细血管管腔变窄或完全阻塞,以致有滤过功能的肾小球数量减少,有效滤过面积也因而减少,导致肾小球滤过率降低,结果出现少尿(每昼夜尿量在100-500ml之间)以致无尿(每昼夜尿量不到100ml)。
表8-1 物质的有效半径和肾小球滤过能力的关系
| 物质 | 分子量 | 有效半径(nm) | 滤过能力 |
| 水 | 18 | 0.10 | 1.0 |
| 钠 | 23 | 0.14 | 1.0 |
| 尿素 | 60 | 0.16 | 1.0 |
| 葡萄糖 | 180 | 0.36 | 1.0 |
| 蔗糖 | 342 | 0.44 | 1.0 |
| 菊粉 | 5500 | 1.48 | 0.98 |
| 肌球蛋白 | 17000 | 1.95 | 0.75 |
| 卵白蛋白 | 43000 | 2.85 | 0.22 |
| 血红蛋白 | 68000 | 3.25 | 0.03 |
| 血浆白蛋白 | 69000 | 3.55 | <0.01 |
滤过能力(filterability)值为1.0表示该物质可自由滤过,0则表示不能滤过
不同物质通过肾小球滤过膜的能力决定于被滤过物质的分子大小及其所带的电荷。表8-1表示被滤过物质的分子量和有效半径对滤过的影响。一般来说,有效半径小于1.8nm的物质,如葡萄糖(分子量180)的有效半径为0.36nm,它可以被完全滤过。有效半径大于3.6nm的大分子物质,如血浆白蛋白(分子量约69000)则几乎完全不能滤过。有效半径介于葡萄糖和白蛋白之间的各种物质,随着有效半径的增加,它们被滤过的量逐渐降低,以上事实提示,滤过膜上存在着大小不同的孔道,小分子物质很容易通过各种大小的孔道,而有效半径较大的物质只能通过较大的孔道,用不同有效半径的中性右旋糖酐分子进行实验,也清楚地说明了被滤过物质的大小对滤过的影响。有效半径小于1.8nm的中性右旋糖酐能自由通过滤过膜,有效半径大于3.6nm的右旋糖酐就完全不能通过。有效半径在1.8-3.6nm的右旋糖酐,其滤过量与有效半径成反比,即随着有效半径增大,滤过量就不断减少(图8-6)。
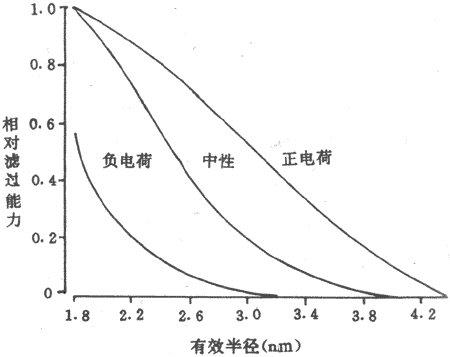
图8-6 不同的有效半径和带不同电荷对右旋糖酐滤过能力的作用
滤过能力的值为1.0,表示自由滤过 0则不能滤过
滤过膜的通性还决定于被滤过物质所带的电荷。用带不同电荷的右旋糖酐进行实验观察到,即使有效半径相同,带正电荷的右旋糖酐较易被滤过,而带负电荷的右旋糖酐则较难通过(图8-6)。血浆白蛋白虽然其有效半径为3.5nm,由于其带负电荷,因此就难于通过滤过膜。
滤过膜的上述特性可由滤过膜的超微结构的特点来说明。滤过膜由三层结构组成(图8-3);①内层是毛细血管的内皮细胞。内皮细胞有上许多直径50-100nm的小孔,称为窗孔(fenestration),它可防止血细胞通过,但对血浆蛋白的滤过可能不起阻留作用。②中间层是非细胞性的基膜,是滤过膜的主要滤过屏障。基膜是由水合凝胶(hydrated gel)构成的微纤维网结构,水和部分溶质可以通过微纤维网的网孔。有人把分离的基膜经特殊染色证明有4-8nm的多角形网孔。微纤维网孔的大小可能决定着分子大小不同的溶质何者可以滤过。③外层是肾小囊的上皮细胞。上皮细胞具有足突,相互交错的足突之间形成裂隙。裂隙上有一层滤过裂隙膜(filtration slit membrane),膜上有直径4-14nm的孔它是滤过的最后一道屏障。通过内、中两层的物质最后将经裂隙膜滤出,裂隙膜在超滤作用中也很重要。
滤过膜各层含有许多带负电荷的物质,主要为糖蛋白。这些带负电荷的物质排斥带带负电荷的血浆蛋白,限制它们的滤过。肾在病理情况下,滤过膜上带负电荷的糖蛋白减少或消失,就会导致带负电荷的血浆蛋白滤过量比正常时明显增加,从而出现蛋白尿。
二、有效滤过压
肾小球滤过作用的动力是有效滤过压。像其它器官组织液生成的机制那样,肾小球有效滤过压=(肾小球毛细血管压+囊内液胶体渗透压)-(血浆胶体渗透压+肾小囊内压)(图8-7)。由于肾小囊内的滤过液中蛋白质浓度较低,其胶体渗透压可忽力略不计。因此,肾小球毛细血管血压是滤出的唯一动力,而血浆胶渗透压和囊内压则是滤出的阻力。有效滤过压=肾小球毛细血管压-(血浆胶体渗透压+肾小囊内压)。皮质肾单位的入球小动脉粗而短,血流阻力较小;出球小动脉细而长,血流阻力较大。因此,肾小球德细血管血压较其它器官的毛细血管血压高。用微穿刺法没得肾小球毛细血管平均值为6.0kPa(45mmHg)(为主动脉平均压的40%左右);用微穿法还发现,由肾小球毛血管的入球端到出球端,血压下降不多,两端的血压几乎相等。肾小囊内压与近曲小管内压力相近。囊内压为1.3kPa(10mmHg)。据测定,在大鼠的肾小球毛细血管入球端的血浆胶体渗透压约为3.3kPa(25mmHg)左右。

图8-7 有效滤过示意图
在入球端,有效滤过压=6.0-(3.3+1.3)=1.4kPa。但肾小球毛细血管内的血浆胶体渗透压不是固定不变的。在血液流经肾小球毛细血管时,由于不断生成滤过液,血液中血浆蛋白浓度就会逐渐增加,血浆胶体渗透压也随之升高。因此,有效滤过压也逐渐下降。当有效滤过压下降到零时,就达到滤过平衡(filtration equilibrium),滤过便停止了(图8-8)。由此可见,不是肾小球毛细血管全段都有滤过作用,只有从入球小动脉端到滤过平衡这一段才有滤过作用。滤过平衡越靠近入球小动脉端,有效滤过的毛细血管长度就越短,有效滤过压和面积就越小,肾小球滤过率就低。相反,滤过平衡越靠近出球小动脉端,有效滤过的毛细血管长度越长,有效滤过压和滤过面积就越大,肾小球滤过率就越高。如果达不到滤过平衡,全段毛细血管都有滤过作用(图8-8)。
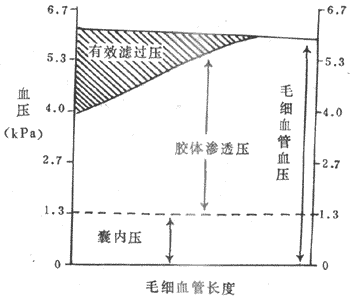
图8-8肾小球毛细血管血压,胶体渗透压和囊内压对肾小球滤过率的作用
三、影响肾小球滤过的因素
滤过膜的通透性和滤过面积的改变对肾小球滤过功能的影响前已述。下面进一步分析肾小球毛细血管血压、血浆胶体渗透压、囊内压和肾血浆流量变化对肾小球滤过功能的影响。
(一)肾小球毛细血管血压
全身动脉血压如有改变,理应影响肾小球毛细血管的血压。由于肾血流量具有自身调节机制,动脉血压变动于10.7?/FONT>24kPa(80-45mmHg)范围内时,肾小球毛细血管血压维持稳定,人而使肾小球滤过率基本保持不变(图8-5)。但当动脉血压降到10.7kPa(80mmHg)以下时,肾小球毛细血管将相应下降,于是有效滤过压降低,肾小球滤过率也减少。当动脉血压降到5.3-6.7kPa(40-50mmHg)以下时,肾小球滤过率将降低到零,因而无尿。在高血压病晚期,入球小动脉由于硬化而缩小,肾小球毛细血管血压可明显降低,于是肾小球滤过率减少而导致少尿。
(二)囊内压
在正常情况下,肾小囊内压是比较稳定的。肾盂或输尿管结石、肿瘤压迫或其他原因引起的输尿管阻塞,都可使肾盂内压显着升高。此时囊内压也将升主,致使有效滤过压降低,肾小球滤过率因此而减少。有些药物如果浓度太高,可在肾小管液的酸性班干部析出结晶;某些疾病时溶血过多,血红蛋白过可堵塞肾小管,这些情况也会导致囊内压升高而影响肾小球滤过。
(三)血浆胶体渗透压
人体血浆胶渗透坟在正常情况下不会有很大变动。但若全身血浆蛋白的浓度明显降低时,血浆胶体渗透压也将降低。此时有效滤过压将升高,肾小球滤过率也随之增加。例如由静脉快速注入生理盐水时,肾小球滤过率将增加,其原因之一可能是血浆胶体渗透压的降低。
(四)肾血浆流量
肾血浆流量对肾小球滤过率有很大影响,主要影响滤过平衡的位置。如果肾轿浆流量加大,肾小球毛细血管内血浆胶体渗透压的上升速度减慢,滤过平衡就靠近出球小动脉端,有效滤过压和滤过面积就增加,肾小球滤过率将随之增加。如果肾血流量进一步增加,血浆胶体渗透压上升速度就进一步减慢,肾小球毛细血管的全长都达不到滤过平衡,全长都有滤过,肾小球滤过率就进一步增加。相反,肾血浆流量减少时,血浆胶体渗透压的上升速度加快,滤过平衡就靠近入球小动脉端,有效滤过压和滤过面积就减少,肾小球滤过率将减少(图8-8)。在严重缺氧、中毒性休克等病理情况下,由于交感神经兴奋,肾血流量和肾血浆流量将显著减少,肾小球滤过率也因而显著减少。
第三节 肾小管与集合管的转运功能
人两肾每天生成的肾小球滤过液达180L,而终尿仅为1.5L。这表明滤过液中约99%的水被肾小管和集合管重吸收,只有约1%被排出体外。不仅如此,滤过液中的葡萄糖已全部被肾小管重吸收回血;钠、尿素告示不同程度地重吸收;肌酐、尿酸和K+等还被肾小管分泌入管腔中。
一、肾小管与集合管的转运方式
肾小管和集合管的转运包括重吸收和分泌。重吸收是指物质从肾小管液中转运至血液中,而分泌是指上皮细胞本身产生的物质或血液中的物质转运至肾小管腔内。肾不球滤过液进入肾小管后称为小管液。
物质通过细胞的转运包括被动转运和主动转运。被动转运是指溶质顺电化学梯度通过肾小管上皮细胞的过程。水的渗透夺之差是水的转运动力。水从渗透压低一侧通过细胞膜进入渗透压高一侧。
主动转运是指溶质逆电化学梯度通过肾小管上皮细胞的过程。主动转运需要消耗能量,根据主动转运过程中能量来源的不同,分为原发性主动转运和继发性主动转运。原发性主动转运(简称为主动转运)所需要消耗的能量由ATP水解直接提供。例如Na+和K+的主动转运是靠细胞膜上的Na+泵水解ATP直接提供能量的。继发性主动转运所需的能量不是直接来自Na+泵。而是来自其他溶质顺电化学梯度转运时释放的。例如一些物质的继发性主动转运的动力直接来自Na+顺电化学梯度转运时释放的能量。释放的之些能量归根到底也是来自Na+泵。由于上皮细胞基侧膜上存在Na+,将细胞内的Na+泵至细胞外,造成细胞内的Na+浓度明显低于细胞外,细胞外K+被泵回细胞内,造成细胞内K+浓度明显高于细胞外,并维持细胞内的负电位。这样,小管液中的Na+便顺电化学梯度通过管腔膜进入细胞,并释放能量提供其他物质的转运。许多物质的转运都与Na+的主动转运相耦联,例如小管液中的葡萄糖、氨基酸、有机酸和CI-等物质的重吸收都与Na+同向转运(cotransport)有关。同向转运是指两种物质与细胞膜上的同向转运体(cotransporter,symporter)特殊蛋白质结合,以相同方向通过细胞膜的转运;又如肾小管细胞分泌H+是与Na+的逆向转运相耦联。逆向转运(antiport)是指两种物质与细胞膜上的逆向转运体(antiport)又称交换体(exchanger)结合,以相反方向通过细胞膜的转运。可见,Na+的主动转运在肾小管上皮细胞的转运中起着关键作用(图8-9)。一个带下电荷和另一个带负电荷的两种物质的同向转动,或电荷相同的两种物质的逆向转运都不会造成小管内外电位改变,这种转运称为电中性转动。如果一个物质是离子,另一个是电中性物质,这种转运就会使小管内外出现电位差,称为生电性转运。如在近球小管,Na+与葡萄糖的同向转运,因葡萄糖是电中性物质,Na+和葡萄糖被重吸收就会造成小管内较小管外带负电位。又如在近球小管的后半段,小管液CI-浓度比管外高,CI-顺浓度差被动重吸收造成管内带正电位。
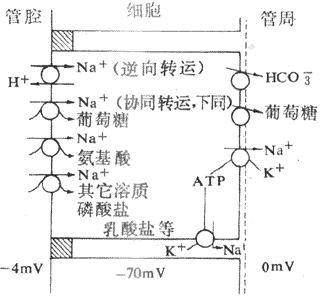
图8-9 Na+转运与其他溶质转运之间的伴联关系
二、各段肾小管和集合管的转运功能
(一)近球小管
肾小球滤过流经近球小管后,滤过液中67%Na+、CI-、K+和水被重吸收,85%的HCO3也被重吸收,葡萄糖、氨基酸全部被重吸收;H+则分泌到肾小管中。近球小管重吸收的关键动力是基侧膜上的Na+泵;许多溶质,包括水的重吸收都与Na+泵的活动有关。
1.Na+、CI-和水的重吸收在近球小管前半段,大部分Na+与葡萄糖,氨基酸同向转运、与H+逆向转运而被主动重吸收;面近球小管后半段,Na+和CI-主要通过细胞旁路而被被动重吸收。水随NaCI等溶质重吸收而被重吸收,因此,该段小管注与血浆渗透压相同,是等渗重吸收。
在近球小管前半段,由于Na+泵的作用,Na+被泵至细胞间隙,使细胞内Na+浓度低,细胞内带负电位。因此,小管液中的Na+和葡萄糖与管腔膜上的同向转运体结合后,Na+顺电化学梯度通过管腔膜的同时,释放的能量将葡萄糖同向转运入细胞内。进入细胞内的Na+即被细胞基侧膜上的Na+泵泵出至细胞间隙,这样,一方面使细胞内Na+的浓度降低,小管液中的Na+-葡萄糖便可不断转运进入细胞内,细胞内的葡萄糖由易化扩散通过细胞基侧膜离开细胞回到血液中;另一方面,使细胞间隙中的Na+浓度升高,渗透压也升高,通过渗透作用,水随之进入细胞间隙。由于细胞间隙在管腔膜侧的紧密连接相对是密闭的,Na+和水进入后就使其中的静水压升高,这一压力可促使Na+和水通过基膜进入相邻的毛细血管而被重吸收,但也可能使部分Na+和水通过紧密连接回漏(back-leak)至小管腔内(图8-10A)。
另一部分的Na+-H+交换而主动重吸收。小管液中的Na+和细胞内的H+与管腔膜上的交换体结合进行逆向转运,使小管液中的Na顺浓度梯度通过管腔膜进入细胞的同时,将细胞内的H+分泌到小管液中;进入细胞内的Na+随即被基侧膜上的Na+泵泵至细胞间隙而主动重吸收。分泌到小管液中的H+将有利于小管液中的HCO3的重吸收。
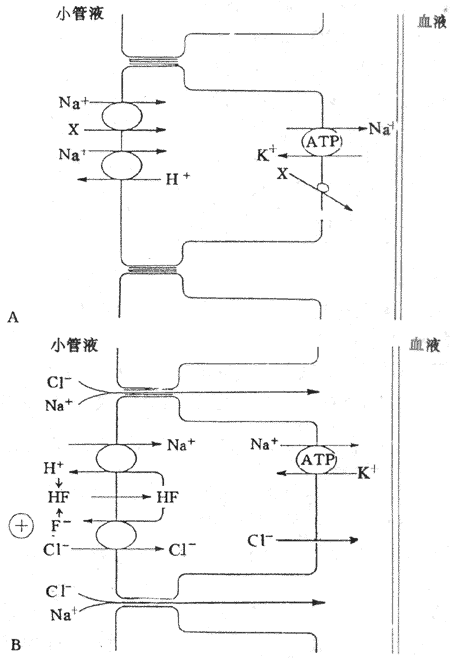
图8-10 近球小管重吸收NaCI的示意图
A:近球小管的前半段 X代表葡萄糖、氨基酸、磷酸盐CI-
B:近球小管的后半段F-代表甲酸盐 HF,甲酸
在近球小管后半段,NaCI是通过细胞旁路和跨上皮细胞两条途径而被重吸收的。小管液进入近球小管后半段时,绝大多数的葡萄糖、氨基酸已被重吸收。由于HCO3重吸收速率明显大于CI-重吸收,CI-留在小管液中,造成近球小管后半段的CI-浓度比管周组织间液高20%-40%。因此,CI-顺浓度梯度经细胞旁路(即通过紧密连接进入细胞间隙)而重吸收回血。由于CI-被动重吸收是生电性的,使小管液中正离子相对较多,造成管内外电位差,管腔内带正电,管外带负电,在这种电位差作用下,Na+顺电位差通过细胞旁路而被动重吸收。CI-通过细胞旁路重吸收是顺浓度梯度进行的,而Na+通过细胞旁路重吸收是顺电位梯度进行的,因此,NaCI是重吸收都是被动的(图8-10B)。
NaCI跨上皮细胞重吸收与H+和甲酸盐(formate,F-)再循环有关,要通过管腔膜上相互耦联的Na+-H+交换和CI—甲酸根交换。在正常肾小管液中含有低浓度甲酸根,通过Na+、H+交换,Na+进入细胞,H+分泌到小管液中,并与小管液中的甲酸根结合形成甲酸(formic acid,HF)。甲酸是脂溶性的,可迅速通过管腔膜进入细胞,在细胞内分解为H+和甲酸根。甲酸根和小管液中的CI-进行逆向转运,结果,CI-进入细胞内并通过基侧膜而被重吸收,而甲酸根则人细胞内进入小管液。细胞内的H+则与小管液中的Na+进行逆向交换,Na+进入细胞,并被Na+泵泵至细胞间隙,然后进入管周毛细血管而被重吸收;H+分泌至小管液,再与小管液中的甲酸根据结合,形成甲酸再进入细胞。因此,H+和甲酸根可再循环使用,CI-和Na+则被重吸收回血。
水的重吸收是被动的,是靠渗透作用而进行的。水重吸收的渗透梯度存在于上皮细胞和细胞间隙之间。这是由于Na+顺电化学梯度通过管腔膜进入细胞后,细胞内的Na+被基侧膜上的Na+泵泵至细胞间隙,使细胞间隙渗透压升高。在渗透作用下,水便不断从小管液进入上皮细胞,并从细胞不断进入细胞间隙,造成细胞间隙静水压升高;加上管周毛细血管内静水压较低,胶体渗透压较高,水便通过周围组织间隙进入毛细血管而被重吸收。
2.HCO3重吸收与H+的分泌 HCO3的重吸收与小管上皮细胞管腔膜上的Na+-H+交换有密切关系。HCO3在血浆中钠盐(NaHCO3)的形式存在,滤过中的NaHCO3滤入囊腔进入肾小管后可解离成Na+和HCO3。通过Na+-H+交换,H+由细胞内分泌到小管液中,Na+进入细胞内,并与细胞内的HCO3一起被转运回血(图8-11)。由于小管液中的HCO3不易通过管腔膜,它与分泌的H+结合生成H2CO2,在碳酸酐酶作用下,H2CO2迅速分解为CO2和水。CO2是高度脂溶性物质,能迅速通过管腔膜进入细胞内,在碳酸酐酶作用下,进入细胞内的CO2与H2O结合生成H2CO3。H2CO3又解离成H+和HCO3。H+通过Na+-H+交换从细胞分泌到小管液中,HCO3则与Na+一起转运回血。因此,肾小管重吸收HCO3是以CO2的形式,而不是直接以HCO3的形式进行的。如果滤过的HCO3超过了分泌的H+,HCO3就不能全部(以CO2形式)被重吸收。由于它不易透过管腔膜,所以余下的便随尿排出体外。可见肾小管上皮细胞分泌1H+就可使1HCO3和1Na+重吸收回血,这在体内的酸碱平衡调节中起到重要作用。乙酰唑胺可抑制碳酸酐酶的活性,因此,用乙酰唑受后,Na+-H+交换就会减少,因而NaHCO3、NaCI和水的排出增加,可引起利尿。由于近球小管的Na+-H+交换,小管液中的HCO3与H+结合并生成CO2,CO2透过管腔膜的速度明显高于CI-的速度。因此,HCO3的重吸收率明显大于CI-的重吸收率。

图8-11 肾小管上皮细胞生成和分泌H+示意图
3.K+的重吸收微穿刺实验表明,肾小球滤过的K+,67%左右在近球小管重吸收回血,而尿中的K+主要是由远曲小管和集合管分泌的。有人认为,近球小管对K+的重吸收是一个主动转运过程。小管液中钾浓度为4mmol/L,大大低于细胞内K+浓度(150mmol/L)。因此在管腔膜处K+重吸收是逆浓度梯度进行的。管腔膜K+主动重吸收的机制尚不清楚。
4.葡萄糖重吸收肾小球滤过液中的葡萄糖浓度与血糖浓度相同,但尿中几乎不含葡萄糖,这说明葡萄糖全部被重吸收因血。微穿刺实验表明,重吸收葡萄糖的部位仅限于近球小管,尤其是在近球小管前半段,其他各段肾小管都没有重吸收葡萄糖的能力。因此,如果在近球小管以后的小管液中仍含有葡萄糖,则尿中将出现葡萄糖。
葡萄糖是不带电荷的物质,它的逆浓度梯度重吸收的,是由Na+继发性主动同向转运而被重吸收的。在兔肾近球小管微灌流实验中观察到,如果灌流液中去掉葡萄糖等有机溶质,则Na+的重吸收率降低;如果灌流液中的Na+全部去掉,则葡萄糖有机溶质的重吸收将完全停止,说明葡萄糖的重吸收与Na+同向转运密切相关。葡萄糖和Na+分别与管腔膜上的同向转运体蛋白的结合位点相结合而进行同向转运(见前述Na+重吸收)。
近球小管对葡萄糖的重吸收有一定限度。当血液中葡萄糖浓度超过160-180mg/100ml时,有一部分肾小管对葡萄糖的吸收已达到极限,尿中开始出现葡萄糖,此时的血糖浓度称为肾糖阈。血糖浓度再继续升高,尿中葡萄糖含量也将随之不断增加;当血糖浓度超过300mg/100ml后,全部肾小管对葡萄糖的吸收均已达到极限,此值即为葡萄糖吸收极限量。此时,尿葡萄糖排出率则随血糖浓度升高而平行增加。人肾的葡萄糖吸收极限量,在体表面积为1.73m2的个体,男性为375mg/min,女性为300mg/min。肾之所以有葡萄糖吸收极限量,可能是由于同向转运体的数目有限的缘故,当所有同向转运体的结合位点都被结合而达饱和时,葡萄糖转运量就无从再增加了。
5.其他物质的重吸收和分泌小管注保的氨基酸的重吸收与葡萄糖的重吸收机制相同,也与Na+同向转运(图8-9)。但是,转运葡萄糖的和转运氨基酸的同向转运体可能不同,也就是说同向转运体具有特异性。此外,HPO4-2、SO4-2的重吸收也也Na+同向转运而进行。正常时进入滤液中的微量蛋白质则通过肾小管上皮细胞吞饮作用而被重吸收。
体内代谢产物和进入体仙的某些物质如青霉素、酚红,大部分的利尿药等,由于与血浆中蛋白结合而不能通过肾小球滤过,它们均在近球小管被主动分泌到小管液中而排出体外。
(二)髓袢
近球小管液流经髓袢过程中,约20%的Na+、CI-、和K+等物质被进一步重吸收。髓袢升支粗段的NaCI重吸收在尿液稀释和浓缩机制中具有重要意义。髓袢升支粗段CI-是逆电化学梯度被上皮细胞重吸收的。微穿刺实验证明,兔髓袢升支粗段管腔内为正电位(+10mV)。在微灌流实验中,如果灌流液中不含K+,则管内的正电位基本消失,CI-重吸收率很低,这说明管腔内正电位与CI-的重吸收和小管液中的K+有密切关系。如果在髓袢升支粗段管周的浸溶液中加入选择性Na+泵抑制剂哇巴因(ouabain)抑制Na+泵后,CI-的转运也受阻,说明Na+泵是CI-重吸收的重要因素。据上述实验,有人提出Na+:2CI-:K+同向转运模式来解释升支NaCI的继发性主动重吸收。该模式认为:①髓袢升支粗段上皮细胞基侧膜上的Na+泵,将Na+由细胞内示向组织间液,使细胞内的Na+下降,造成管腔内与细胞内Na+有明显的浓度梯度;②Na+与管腔膜上同向转运体结合,形成Na+:2CI-:K+同向转运体复合物,Na+顺电化学梯度将2CI-和K+一起同向转运至细胞;③进入细胞内的Na+、2CI-和K+的去向各不相同:Na+由Na+泵泵至组织间液,2CI-由于浓度梯度经管周膜上CI-通道进入组织间液,而K+则顺浓度梯度经管腔膜而返回管腔内,再与同向转运体结合,继续参与Na+:2CI-K+的同向转运,循环使用;④由于2CI-进入组织间液,K+返回管腔内,这就导致管腔内出现正电位;⑤由于管腔内正电位,使管腔液中的Na+等正离子顺电位差从细胞旁路进入组织间液,这是不耗能的Na+被动重吸收。从这个模式说明,通过Na+泵的活动,继发性主动重吸收了2CI-,同时伴有2Na+的重吸收,其中1Na+是主动重吸收,另1Na+通过细胞旁路而被动重吸收,这样为Na+的重吸收节约了50%能量消耗(图8-12)。髓袢升支粗段对水的通透性很低,水不被重吸收而留在小管内。由于NaCI被上皮细胞重吸收至组织间液,因此造成小管液低渗,组织间液高渗。这种水和盐重吸收的分离,有利于尿液的浓缩和稀释。Na+:2CI-:K+同向转运对速尿,利尿酸等利尿剂很敏感。它们与同向转运体结合后,可抑制其转运功能,管腔内正电位消失,NaCI的重吸收受抑制,从而干扰尿的浓缩机制,导致利尿。
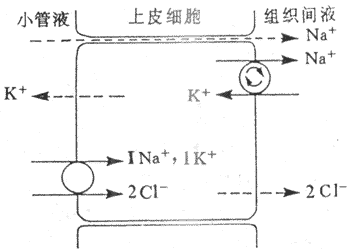
图8-12 髓袢升支粗段继发性主动吸收CI-的示意图
(三)远球小管和集合管
在远曲小管和集合管,重吸收大约12%滤过的Na+和CI-,分泌不同量的K+和H+,重吸收不同量的水。水、NaCI的重吸收以及K+和H+的分泌可根据机体贴的水、盐平衡状况来进行调节。如相机缺水或缺盐时,远曲小管和集合管可增加水、盐的重吸收;当机体水、盐过剩时,则水、盐重吸收明显减少,水和盐从尿排出增加。因此,远曲小管和集合管对水和盐的转运是可被调节的。水的重吸收主要受抗利尿激素调节,而Na+和K+的转运主要受醛固酮调节。
远曲小管和集合管上皮细胞间隙的紧密连接对小离子如Na+、K+和CI-等的通透性低,这些离子不易通过紧密连接回漏至小管腔内,因此,所能建立起来的管内外离子浓度梯度和电位梯度大。在远曲小管初段,对水的通透性很低,但仍主动重吸收NaCI,继续产生低渗小管液。Na+在远曲小和集合管的重吸收是逆较大的电化学梯度进行的,是主动重吸收过程。这可能与远曲小管的Na+泵在肾单位中活性最高有关。有人认为在远曲小管初段的小管液中,Na+是通过Na+-CI-同向转运进入细胞的,然后由Na+泵将Na+泵出细胞而主动重吸收回血(图8-13A)。NaCI同向转运体可被噻嗪类利尿药所抑制。
远曲小管后段和集合管含有两类细胞,即主细胞和闰细胞。主细胞重吸收Na+和水,分泌K+,闰细胞则主要分泌H+,主细胞重吸收Na+主要通过管腔膜上的Na+通道。管腔内的Na+顺电化学梯度通过管膜上的Na+通道进入细胞,然后,由Na+泵泵至细胞间液而被重吸收(图8-13B)。
K+的分泌 尿中K+的排泄量视K+的摄入量而定,高钾饮食可排出大量的钾,低钾饮食则尿中排钾量少,使机体的钾摄入量与排出量保持平衡,维持机体K+浓度的相对恒定。
K+分泌的动力包括:①在远曲小管和集合管的小管液中,Na+通过主细胞的管腔膜上的Na+通道进入细胞,然后,由基侧膜上的Na+泵将细胞内的Na+泵至细胞间隙而被重吸收,因而是生电性的,使管腔内带负电位(-10?40mV)。这种电位梯度也成为K+从细胞分泌至管腔的动力;②在远曲小管后段和集合管的主细胞内的K+浓度明显高于小管液中的K+浓度,K+便顺浓度梯度从细胞内通过管腔膜上的K+通道进入小管液;③Na+进入主细胞后,可刺激基侧膜上的Na+,使更多的K+从细胞外液中泵入细胞内,提高细胞内K+浓度,增加细胞内和小管液之间的K+浓度梯度,从而促进K+分泌,因此,K+的分泌与Na+的重吸收有密切关系。(图8-13B)。

图8-13 远球小管和集合管重吸收NaCI、分泌K+和H+的示意图
A:远曲小管初段 B:远曲小管后段和集合管
H+的分泌除了近球小管细胞通过Na+-H+交换分泌H+,促进NaHCO3重吸收外,远曲小管和集合管的闰细胞也可分泌H+。H+的分泌是一个逆电化学梯度进行的主动转运过程。有人认为管腔膜上有H+泵,能将细胞内的H+历史意义入小管腔内。细胞内的CO2和H2O在碳酸酐酶催化作用下生成的H+和HCO3,H+由H+泵泵至小管液,HCO3则通过基侧膜回到血液中,因而H+分泌和HCO3的重吸收与酸碱平衡的调节有关(图8-13B)。闰细胞分泌的H+与HPO4-2结合形成H2PO4,这是可滴定酸;分泌的H+可与上皮细胞分泌的NH3结合,形成NH4+。可滴定酸和NH4+都不易透过管腔膜进入细胞而留在小管液中。因此,它们是尿液酸碱度的决定因素。
NH3的分泌远曲小管和集合管的上皮细胞在代谢过程中不断地生成NH3,这些NH3主要由谷氨酰胺脱氨而来。NH3具有脂溶性,能通过细胞膜向小管液周围组织间液和小管液自由扩散,扩散量取决于两种液体的pH值。小管液的pH较低(H+浓度较高),所以NH3较易向小管液中扩散。分泌的NH3能与小管液中的H+结合并生成NH4-,小管液中NH3浓度因而下降,于是管腔膜两侧形成了NH3浓度梯度,此浓度梯度又加速了NH3向小管液中扩散。由此可见,NH3的分泌与H+的分泌密切相关;H+分泌增加促使NH3分泌增多。NH3与H+结合并生成NH4-后,可进一步与小管液中的强酸盐(如NaCI等)的负离子结合,生成酸性铵盐(NH4CI等)并随尿排出。强酸盐的正离子(如Na+)则与H+交换而进入肾小管细胞,然后和细胞内HCO3-一起被转运回血。所以,肾小管细胞分泌NH3,不仅由于铵盐形成而促进了排H+,而且也促进了NaHCO3的重吸收。
第四节 尿液的浓缩和稀释
尿的渗透浓度可由于体内缺水或水过剩等不同情况而出现大幅度的变动。当体内缺水时,机体将排出渗透浓度明显高于血浆渗透浓度的主渗尿,即尿被浓缩。而体内水过剩时,将排出渗透浓度低于血浆渗透浓度的低渗尿。正常人尿液的渗透浓度可在约50-1200mOsm/kgH2O之间波动。所以,根据尿的渗透浓度可以了解肾的渗透浓度和稀释能力。肾的浓缩和稀释能力,以维持体液平衡和渗透压恒定中有极为重要的作用。
一、尿液的稀释
尿液的稀释是由于小管液的溶质被重吸收而水不易被重吸收造成的。这种情况主要发生在髓袢升支粗段。前已述及,髓袢升支粗段能主动重吸收Na+和CI-(图8-12),而对水通透,故水不被重吸收,造成髓袢升支粗段小管液为低渗。在体内水过剩而抗利尿激素释放被抑制时,集合管对水的通透性非常低。因此,髓袢升支的小管液流经远曲小管和集合管时,NaCI继续重吸收,使小管液渗透浓度进一步下降。可降低至50mOsm/kgH2O,形成低渗尿,造成尿液的稀释。如果抗利尿激素完全缺乏时,如严重尿崩症患者,每天可排出高达20L的低渗尿,相当于肾小球滤过率的10%。
二、尿液的浓缩
尿液的浓缩是由于小管液中的水被重吸收而溶质仍留在小管液中造成的。水重吸收的动力来自肾髓质渗透梯度的建立,即髓质渗透浓度从髓质外层向乳头部深入而不断升高。用冰点降低法测定鼠肾的渗透浓度观察到肾皮质部的组织间液(包括细胞内液和细胞外液)的渗透浓度与血液渗透浓度之比为1.0,说明皮质部组织间液与血浆是等渗的。而髓质部组织间液与血浆的渗透浓度之比,随着由髓质外层向乳头部深入而逐渐升主,分别为2.0、3.0、4.0(图8-14)。这表明肾髓质的渗透浓度由外向内逐步升高,具有明确的渗透梯度。在抗利尿激素存在时,远曲小管和集合管对水通透性增加,小管液从外髓集合管向内髓集合管流动时,由于渗透作用,水便不断进入高渗的组织间液,使小管液不断被浓缩而变成高渗液,最后尿液的渗透浓度可高达120mOsm/kgH2O,形成浓缩尿。可见髓质的渗透梯度是建立就成为浓缩尿的必要条件。髓袢是形成髓质渗透梯度的重要结构,只有具有髓袢的肾才能形成浓缩尿,髓袢愈长,浓缩能力就愈强。例如沙鼠的肾髓质内层特别厚,它的肾能产生20倍于血浆渗透浓度的高渗尿。猪的髓袢较短,只能产生1.5倍于血浆渗透浓度的尿液。人的髓袢具有中等长度,最多能产生4-5倍于血浆渗透浓度的高渗尿。
髓质渗透梯度是如何形成的?有人用肾小管各段对水和溶质通透性不同(表8-2)和逆流倍增现象来解释。

8-14图 肾髓质渗透压梯度示意图
线条越密,表示渗透压越高
表8-2 兔肾小管不同部分的通透性
| 肾小管部分 | 水 | Na+ | 尿素 |
| 髓袢升支粗段 | 不易通透 | Na+主动重吸收 | 不易通透 |
|
| CI-继发性主动重吸收 |
|
| 髓袢升支细段 | 不易通透 | 易通透 | 中等通透 |
| 髓袢降支细段 | 易通透 | 不易通透 | 不易通透 |
| 远曲小管 | 有ADH时水易通透 | Na+主动重吸收 | 不易通透 |
| 集合管 | 有ADH时水易通透 | Na+主动重吸收 | 皮质稳步髓部不易通透 |
ADH为抗利尿激素*不同动物,通透性不一样
物理学中逆流的涵意是指两个并列的管道,其中液体流动的方向相反,如图8-15所示,甲管中液体向下流,乙管中液体向上流。如果甲乙两管下端是连通的,而且两管间的隔膜容许液体中的溶质或热能在两管间交换,便构成了逆流系统。在逆流系统中,由于管壁通透性和管道周围环境的作用,就会产生逆流倍增现象。
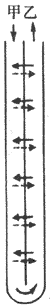
图8-15 逆流系统示意图
甲管内液体向下流 乙管内液体向上流
逆流倍增现象可要根据图8-16的模型来理解。模型中含有钠盐的液体从甲管流进,通过管下端的弯曲部分又折返流入乙管,然后从乙管反向流出,构成逆流系统。溶液流动时,由于M1膜能主动将Na+由乙管泵入甲管,而M1膜对水的通透性又很低,因此,甲管中深夜在向下流动过程中将不断接受由乙管泵入的Na+,于是Na+的浓度不断增加(倍增)。结果甲管中溶液自上而下的渗透浓度会越来越高,到甲管下端的弯曲部分时Na+深夜逐渐下降,渗透浓度也相应下降。这样,不论是甲管还是乙管,从上而下来比较,溶液的渗透浓度均逐渐升高,即出现了逆流倍增现象,形成了渗透梯度。如果有渗透浓度较低的溶液从丙管向下流动,而且M2膜对水不能通透,对溶质不通透,水将因渗透作用而进入乙管。这样丙管内溶质的深夜将逐渐增加;从丙管下端流出的液体成了高渗溶液。

图8-16 逆流倍增作用模型
甲管内液体向下流 乙田径赛内液体向上流丙管内液体向下流 M1膜能将液体
中Na+由乙管泵入甲管,且对水不易通透M2膜对水易通透
髓袢、集合管的结构排列与上述的逆流倍增的模型很相似。这对理解尿的浓缩机制是有帮助的。
在外髓部,由于髓袢升支粗段能主动重吸收Na+和CI-(图8-17),而对水不通透,故升支粗段内小管液向皮质方向流动时,管内NaCI浓度逐渐降低,小管液渗透浓度逐渐下降;而升支粗段外围组织产液则变成高渗。髓袢升支粗段位于外髓部,故外髓部的渗透梯度主要是由升支粗段NaCI的重吸收所形成。愈靠近皮质部,渗透浓度越低;愈靠近内髓部,渗透浓度越高。
在内髓部,渗透梯度的形成与尿素的丙循环和NaCI重吸收有密切关系。①远曲小管及皮质部和外髓部的集合管对尿素不易通透,但小管液流经远曲小管及皮质部和外髓部的集合管时,在抗利尿激素作用下,对水通透性增加,由于外髓部高渗,水被重吸收,所以小管液中尿素的浓度逐渐升高。②当小管液进入内髓部集合管时,由于管壁对尿素的通透性增大,小管液中尿素就顺浓度梯度通过管壁向内髓部组织间液扩散,造成了内髓部组织间液中尿素浓度的增高,渗透浓度因之而长高。③髓袢降支细段对尿素不易通透,而对水则易通透,所以在渗透压的作用下,水被“抽吸”出来,从降支细段进入内髓部组织间液。由于降支细段对Na+不易通透,小管液将被浓缩,于是其中的NaCI浓度愈来愈高,渗透浓度不断升高。④当小管液绕过髓袢顶端折返流入升支细段时,它同组织间注NaCI浓度梯度明显地建立起来。由于升支细段对Na+易通透,Na+将顺浓度梯度而被动扩散至内髓部组织间液,从而进一步提高了内髓部组织间液的渗透浓度。由此看来,内髓部组织间液的渗透浓度,是由内髓部集合管扩散出来的尿素以及髓袢升支细段扩散出来的NaCI两个因素造成的。⑤小管液在升支细段流动过程中,由于NaCI扩散到组织间液,而且该管壁又对水不易通透,所以造成了管内NaCI浓度逐渐降低,渗透浓度也逐渐降低,这样,降支细段与升支细段就构成了一个逆流倍增系统,使内髓部组织间液形成了渗透梯度。⑥尿素是可以再循环的。因为升支细胞对尿素具有中等的通透性,所以从内髓部集合管扩散到组织间液的尿素可以进入升支细段,而后流过升支粗段、远曲小管、皮质部和外髓部集合管,又回到内髓部集合管外再扩散到内髓部组织间液,这样就形成了尿素的丙循环(图8-17)。
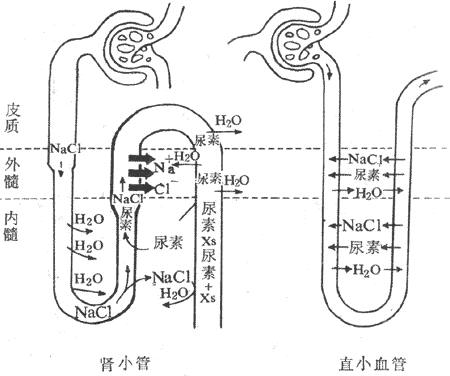
图8-17 尿浓缩机制示意图
粗箭头表示升支粗段主动重吸收Na+和CI-。粗线表示髓袢升支粗段和远曲小管前段对水不通透。字体大小表示溶质深夜。XS表示未被重吸收的溶质
从髓质渗透梯度形成全过程来看,髓袢升支粗段对Na+和CI-的主动重吸收是髓质渗透梯度建立的主要动力。而尿素和NaCI是建立髓质渗透梯度的主要溶质。
三、直小血管在保持肾髓质高渗中的作用
直小血管的功能可用逆流交换现象来理解。图8-18是逆流交换的示意图。在图A中,U形管的升、降支之间不能进行热量交换,所以降支中的冷水在流到热源以前得不到加温,升支中的水温在离开热源以后也不能降低。这样,冷水流过U形管的升、降支之间能够交换热量,所以降支中的冷水在进入热源以前就被升支管壁透过来的热量所加温,而升支中的水则因热量不断透入降支而降温。这样,冷水流过U形管时,从热源带走的热量就很有限,所在热源损失掉的热量也很少。
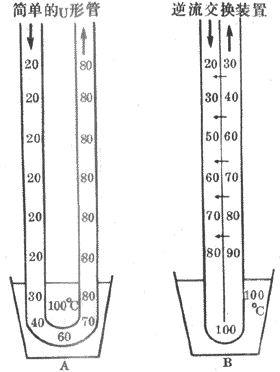
图8-18 逆流交换作用的简单物理模型示意图
下如前述,通过肾小管上述的逆流作用,不断有溶质(NaCI和尿素)进入髓制裁组织间液形成渗透梯度,也不断有水被肾小管和集合管重吸收至组织间液。因此,必须把组织间液中多余的溶质和水被除去才能保持髓质渗透梯度。通过直小血管的逆流交换作用就能保持髓质渗透梯度。直小血管的降支和长支是并行的细血管,这咱结构就是逆流系统。在直小血管降支进入髓质的入口处,其血浆渗透浓度约为300mOsm/kgH2O。由于直小血管对溶质和水的通透性高,当它在向髓质深部下行过程中,周围组织间液中的溶质就会顺浓度梯度不断扩散到直小血管降支中,而其中的水则渗出到组织间液,使血管中的血浆渗透浓度与组织间液达到平衡。因此,愈向内髓部深入,降支血管中的溶质浓度愈高。在折返处,其渗透浓度可高达120mOsmkgH2O。如果直小血管降支此时离开髓质,就会把从进入直小血管降支中的大量溶质流回循环系统,而从直小血管内出来的水保留在组织间液。这样,髓质渗透梯度就不能维持。由于直小血管是逆流系统。因此,当直小血管升支从髓质深部返回外髓部时,血管内的溶质浓度比同一水平组织间液的高,溶质又逐渐扩散回组织间液,并且可以再进入降支,这是一个逆流交换过程。因此当直小血管升支离开外髓部时,只把多余的溶质带回循环中。此外,通过渗透作用,组织间液中的水不断进入直小血管升支,又把组织间液中多余的水随血流返回循环。这样就维持了肾髓质的渗透梯度(图8-17)。
第五节 肾尿生成的调节
尿的生成有赖于肾小球的滤过作用和肾小管、集合管的重吸收和分泌作用。因此,机体对尿的生成的调节也就是通过对滤过作用和重吸收、分泌作用的调节来实现的。肾小球滤过作用的调节在前文已述,本节主要论述肾小管和集合管重吸收和分泌的调节。肾小管和集合管功能的调节包括肾内自身调节和神经、体液调节。
一、肾内自身调节
肾内自身调节包括小管液中溶质浓度的影响、球-管平衡和管-球反馈等。
(一)小管液中溶质的浓度
小管液中溶质所呈现的渗透压,是对抗肾小管重吸收水分的力量。如果小管液溶质浓度很高,渗透压很大,就会妨碍肾小管特别是近球小管对水的重吸收,小管液中的Na+被稀释而浓度下降,小管液中与细胞内的Na+浓度差变小,Na+重吸收减少,因此,不仅尿量增多,NaCI排出也增多。例如糖尿病患者的多尿,就是由于小管液中葡萄糖含量增多,肾小管不能将葡萄糖完全重吸收回血,小管液渗透压因而增高,结果妨碍了水和NaCI的重吸收所造成的。临床上有时给病人使用肾小球滤过而又不被肾小管重吸收的物质,如甘露醇等,利用它来提高小管液中溶质的浓度,借以达到利尿和消除水肿的目的。这种利尿方式称为渗透性利尿。
(二)球-管平衡
近球小管对溶质和水的重吸收量不是固定不变的,而是随肾小于滤过率的变动而发生变化。肾小球滤过率增大,滤液中的Na+和水的总含量增加。近球小管对Na+和水的重吸收率也提高;反之,肾小球滤过率减小,滤液中的Na+和水的总含量也减少,近球小管的Na+的水的重吸收率也相应地降低。实验说明,不论肾小于滤过率或增或减,近球小管是定比重吸收(constant fraction reabsorption)的,即近球小管的重吸收率始终占肾小球滤过率的65%-70%左右(即重吸收百分率为65%-70%)。这种现象称为于-管平衡(glomerulotubular balance)。球管平衡的生理意义在于使尿中排出的溶质和水不致因肾小管滤过率的增减而出现大幅度的变动。例如,在正常情况下,肾小球滤过率为125ml/min,近球小管的重吸收率为87.5ml/min(占70%)。流到肾小管远侧部分的量为37.5ml/min。如果滤过率增加到150ml/min,则近球小管的重吸收率变为105ml/min(仍占70%),而流到肾小管远侧部分的量为45ml/min。这几个数字表明,此时滤过率虽然增加了25ml/min,但流到肾小管远见侧部分的量仅增加7.5ml/min。而且在这种情况下,远侧部分的重吸收也有增加,因此尿量的变化是不大的。同样,滤过率减少到100ml/min,近球小管的重吸收率为70ml/min(仍占70%),流到肾小管远侧部分的量为30ml/min。此时的滤过率虽然减少了25ml/min,但流到肾小管远侧部分的量仅减少了7.5ml/min;而且在这种情况下远侧部分的重吸收也要减少,因此尿量的变化仍然不大。近球小管对Na+也是定比重吸收,即重吸收量为滤过量的65%-70%。如果近球小管对Na+重吸收的总量是固定不变的话,根据测算,肾小球滤过率仅增加2ml/min,Na+的排出量就会比原来的增加约2倍;肾小球滤过率减少2ml/min,尿中就不含Na+,可见球管平衡具有重要的生理意义。
定比重吸收的机制与管周毛细血管血压和胶体渗透压改变有关。比如,在肾血流量不变的前提下,当肾小球滤过率增加时,进入近球小管旁毛细血管的血液量就会减少,,血浆蛋白的浓度相对地增高,此时毛细血管内血压下降而胶体渗透压升高。在这种情况下,小管旁组织间液就加速进入毛细血管,组织间液内静水压因之下降,组织间液内静水压下降使得小管细胞间隙内的Na+和水加速通过基膜而进入小管旁的组织间隙;并且通过紧密连接回流至肾小管腔内的回漏量因此而减少,最后导致Na+和水重吸收量增加。这样,重吸收仍可达到肾小球滤过率的65%-70%。肾小球滤过率如果减少,便发生相反的变化,重吸收百分率仍能保持65%-70%。
球-管平衡在某些情况下可能被打乱。例如,渗透性利尿时,近球小管重吸收率减少,而肾小球滤过率不受影响,这时重吸收百分率就会小于65%-70%,尿量和尿中的NaCI排出量明显增多。
目前认为球-管平衡障碍与临床上见到的某些水肿的形成机制有关。例如在充血性心力衰竭时,肾灌注压和血流量可明显下降。但由于出球小动脉发生代偿性收缩,所以肾小球滤过率仍能保持水平。因此滤过分数将变大。此时近球小管旁毛细血管血压下降而血浆胶体渗透压增高。如上所述,这将导致Na+和水的重吸收增加,重吸收百分率将超过65%-70%。于是因体体内钠盐潴留和细胞外液量增多而发生水肿。
(三)管-球反馈
管-球反馈是肾血流量和肾小球滤过率自身调节的重要机制之一。当肾血流量和肾小球滤过率增加时,到达远曲小管致密斑的小管液的流量增加,致密班发生信息,使肾血流量和肾小球滤过率恢复至正常。相反,肾血流量和肾小球滤过率减少时,流经致密斑的小管液流量就下降,致密斑发生信息,使肾血流量和肾小球滤过率增加至正常水平。这种小管液流量变化影响肾血流量和肾小球滤过率的现象称为管-球反馈(tubuloglomerular feed back)。有人认为致密斑主要感受小管液中的NaCI含量改变而不是小管液的流量。一般来说,肾小管液流量与NaCI含量成正比。致密斑发在管-球环节中起重要的传感器(sensor)作用。致密斑与入球小动脉和出球小动脉相邻。致密斑发出的信息通过某种途径影响入球小动脉的口径,从而影响肾血流量和肾小球滤过率。当肾血流量增加时,肾小球滤过率也增加,流经远曲小管的小管液量也增加,致密斑部位NaCI含量升高,致密斑发出信息刺激颗粒细胞释放肾素,导致局部生成血管紧张素Ⅱ,血管紧张素Ⅱ引起入球小动脉收缩,口径缩小,阻力增加,从而使肾血流量和肾小球滤过率恢复至原来水平。相反,当肾血流量减少时,肾小球滤过率下降,流经远曲小管的小管液流量减少,颗粒细胞释放肾素减少,血管紧张至少Ⅱ生成减少,入球小动脉收缩变弱,口径变粗,阻力减少,肾血流量恢复至原有水平。此外,肾内产生的前列腺素、腺苷和儿茶酚胺等也参与管-球反馈。
二、神经和体液调节
(一)交感神经系统
肾交感神经兴奋通过下列作用影响尿生成:①入球小动脉和出球小动脉收缩,而前者血管收缩比后者更明显,因此,肾小球毛细血管的血浆流量减少和肾小球毛细血管的血压下降,肾小球的有效滤过压下降,肾小球滤过率减少;②刺激近球小体中的颗粒细胞释放肾素,导致循环中的血管紧张素Ⅱ和醛固酮含量增加,增加肾小管对NaCI和水的重吸收;③增加近球小管和髓袢皮皮细胞重吸收Na+、CI-和水。微穿刺表明,低频率低强度电刺激肾交感神经,在不改变肾小球滤过率的情况下,可增加近球小管和髓袢对Na+、CI-和水的重吸收。这种作用可被α1肾上腺素受体拮抗剂所阻断。这些结果表明,肾交感神经兴奋时其末稍释放去甲肾上腺素。作用于近球小管和髓袢细胞膜上的α1肾上腺素能受体,增加Na+、CI-和水的重吸收。抑制肾交感神经活动则有相反的作用。
(二)抗利尿激素
抗利尿激素(antidiuretic hormone,ADH)又称血管升压素(vasopressin,AVP),是由9个氨基酸残基组成的小肽,它是下丘脑的视上核和室旁核的神经元分泌的一种激素。它在细胞体中合成,经下丘脑-垂体束被运输到神经垂体然后释放出来。它的作用主要是提高远曲小管和集合管上皮细胞对水的通透性,从而增加水的重吸收,使尿液浓缩,尿量减少(抗利尿)。此外,抗利尿激素也能增加髓袢升支粗段对NaCI的主动重吸收和内髓部集合管对尿素的通透性,从而增加髓质组织间液的溶质浓度,提高髓质组织间液的渗透浓度,有利于尿注浓缩(见尿液浓缩和稀释)。
抗利尿激素与远曲小管和集合管上皮细胞管周膜上的V2受体结合后,激活膜内的腺甘酸化酶,使上皮细胞中cAMP的生成增加;cAMP生成增加激活上皮细胞中的蛋白激酶,蛋白激酶的激活,使位于管腔膜附近的含有水通道的小泡镶嵌在管腔膜上,增加管腔膜上的水通道,从而增加水的通透性。当抗利尿激素缺乏时,管腔膜上的水通道可在细胞膜的衣被凹陷处集中,后者形成吞饮小泡进入胞浆,称为内移(internalization)。因此,管腔膜上的水通道消失,对水就不通透。这咱含水通道的小泡镶嵌在管腔膜或从管腔膜进入细胞内,就可调节管腔内膜对水的通透性(图8-19)。基侧膜则对水可自由通过,因此,水通过管腔膜进入细胞后自由通过基侧膜进入毛细血管而被重吸收。
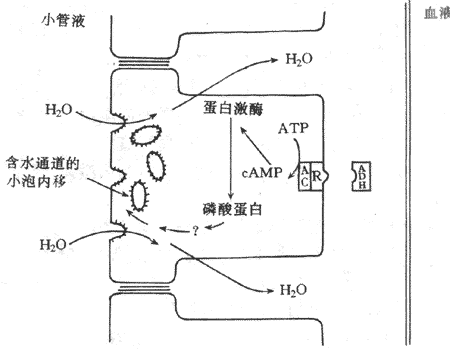
图8-19 抗利尿激素的作用机制示意图
调节抗利尿激素的主要因素是血浆晶体渗透压和循环血量、动脉血压。血浆晶体渗透压的改变可明显影响抗利尿激素的分泌。大量发汗。严重呕吐或腹泻等情况使机体失水时,血浆晶体渗透压升高,可引起抗利尿激素分泌增多,使肾对水的重吸收活动明显增强,导致尿液浓缩和尿量减少。相反,大量饮清水后,尿液被稀释,尿量增加,从而使机体内多余的水排出体外。例如,正常人一次饮用100ml清水后,约过半小时,尿量就开始增加,到第一小时末,尿量可达最高值;随后尿量减少,2-3小时后尿量恢复到原来水平。如果饮用的是等渗盐水(0.9NaCI溶液),则排尿量不出现饮清水后那样的变化(图8-20)。这种大量饮用清水后引起尿量增多的现象,称为水利尿,它是临床上用来检测肾稀释能力的一种常用的试验。循环血量的改变,能反射性地影响抗利尿激素的释放。血量过多时,左心房被扩张,刺激了容量感受器,传入冲动经迷走神经传入中枢,抑制了下丘脑-垂体后叶系统释放抗利尿激素,从而引起利尿,由于排出了过剩的水分,正常血量因而得到恢复。血量减少时,发生相反的变化。动脉血压升高,刺激颈动脉窦压力感受器,可反射性地抑制抗利尿激素的释放。此外,心房尿钠肽可抑制抗利尿激素分泌,血管紧张素Ⅱ则可刺激其分泌。
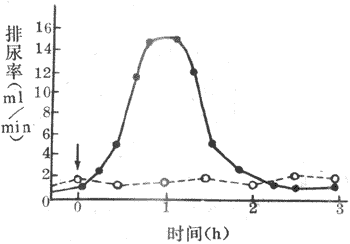
图8-20一次饮一升清水(实线)和饮一升等渗盐水
(0.9NaCI溶液)(虚线)后的利尿率箭头表示饮水时间
(三)肾素-血管紧张素-醛固酮系统
肾素主要是由近球小体中的颗粒细胞分泌的。它是一种蛋白水解酶,能催化血浆中的血管紧张素原使之生成血管紧张素壹I(十肽)。血液和组织中,特别是肺组织中有血管紧张素转换酶,转换酶可使血管紧张素I降解,生成血管紧张素Ⅱ(八肽)。血管紧张素Ⅱ可刺激肾上腺皮质球状带合成和分泌醛固酮。
肾素的分泌受多方面因素的调节。目前认为,肾内有两种感受器与肾素分泌的调节有关。一是入球小动脉处的牵张感受器,另一是致密斑感受器。当动脉血压下降,循环轿量减少时,肾内入球小动脉的压力也下降,血流量减少,于是对小动脉壁的牵张刺激减弱,这便激活了牵张感受器,肾素释放量因此而增加;同时,由于入球小动脉的压力降低和血流量减少,于是激活了致密斑感受器,肾素释放量也可增加。据推想,在近球小体的颗粒细胞和致密斑之间有一种特殊的联系。当两者接触增加时,肾素分泌便减少,而两者接触减少时,则肾素分泌增加。入球小动脉的压力下降,血流量减少时,血管口径缩小,于是颗粒细胞和致密班的接触减少,此时肾素分泌增加;当致密斑处Na+量和小管液量减少时,肾小管口径缩小,两者的接触减少,肾素分泌增加。但这种推想尚缺乏实验证据。此外,颗粒细胞受交感神经支配,肾交感神经兴奋时(如循环血量减少)能引致肾素的释放量增加。肾上腺素和去甲肾上腺素也可直接刺激颗粒细胞,促使肾素释放增加。
1.血管紧张素Ⅱ对尿生成的调节包括:①刺激醛固酮的合成和分泌;醛固酮可调节远曲小管和集合管上皮细胞的Na+和K+转运;②可直接刺激近球小管对NaCI的重吸收,使尿中排出的NaCI减少;③刺激垂体后叶释放抗利尿激素,因而增加远曲小管和集合管对水的重吸收,使尿量减少。
2. 醛固酮对尿生成的调节醛固酮是肾上腺皮质球状带分泌的一种激素。它对肾的作用是促进远曲小管和集合管的主细胞重吸收Na+,同时促进K+的排出,所以醛固酮有保Na+排K+作用。
醛固酮进入远曲小管和集合管的上皮细胞后,与胞浆受体结合,形成激素-受体复合物;后者通过核膜,与核中的DNA特异性结合位点相互作用,调节特异性mRNA转录,最后合成多种的醛固酮诱导蛋白(aldosterone-induced protein)。醛固酮诱导蛋白可能是:①管腔膜的Na+通道蛋白,从而增加管腔的Na+通道数量;②线粒体中合成的ATP的酶,增加ATP的生成,为上皮细胞活动(Na+泵)提供更多的能量;③基侧膜的Na+泵,增加Na+泵的活性,促进细胞内的Na+泵回血液和K+进入细胞,提高细胞内的K+浓度,有利于K+分泌(图8-21);由于Na+重吸收增加,造成了小管腔内的负电位,有利于K+的分泌和CI-的重吸收。结果,在醛固酮的作用下,远曲小管和集合管对Na+和集合管对Na+的重吸收增强的同时,CI-和水的重吸收增加,导致细胞外液量增多;K+的分泌量增加。

图8-21 醛固酮作用机制的示意图
醛固酮的分泌除了受血管紧张素调节外,血K+浓度升高和血Na+浓度降低,可直接刺激肾上腺皮质球状带增加醛固酮的分泌,导致保Na+排K+,从而维持了血K+和血Na+浓度的平衡;反之,血K+浓度降低,或血Na+浓度升高,则醛固酮分泌减少。醛固酮的分泌对血K+浓度升高十分敏感,血K+仅增加0.5-1.0mmol/L就能引起醛固酮分泌。而血Na+浓度必须降低很多才能引起同样的反应。
(四)心房利尿钠肽
心房利尿钠肽(atrial natriuretic pepitde,ANP)是心房肌合成的激素。循环中的心房利尿钠肽是由28个氨基酸残基组成的。它有明显的促进NaCI和水的排出作用。其作用机制可能包括:①抑制集合管对NaCI的重吸收。心房利尿钠肽与集合管上皮细胞基侧膜上的心房利尿钠肽受体结合,激活了鸟苷酸化酶,造成细胞内cGMP含量增加,后者使管腔膜上的Na+通道关闭,抑制Na+重吸收,增加NaCI的排出;②使出球小动脉、尤其是入球小动脉舒张,增加肾血浆流量和肾小球滤过率③抑制肾素的分泌;④抑制醛固酮的分泌;⑤抑制抗利尿激素的分泌。
第六节 清除率
一、清除率的概念和计算方法
清除率是一个抽象的概念,它把一肾在一定时间内排出的物质的量,同当时该物质在血浆中浓度联系起来,因而能更她地说明肾排出某物质的能力。为了讲清楚它的涵意,现举例说明。某甲每分钟尿量为1ml(V=1ml/min),尿中某物质的浓度(U)为100mg/100ml,血浆中该物质的浓度(P)为1mg/100ml。某乙每分钟尿量为0.8ml(V=0.8ml/min),尿中该物质的浓度(U)为50mg/100ml,血浆中该物质的浓度(P)为0.32mg/100ml。经计算,甲肾的该物质排泄量应为U×V=0.8ml/min×50mg/100ml=0.4mg/min。从每分钟该物质排泄量的多少来看,甲肾的功能似乎比乙肾好,其实则不然。因为甲血浆中该物质的浓度比乙肾血浆中的高三倍多,甲肾清除血浆中该物质比较容易,而乙肾则比较困难。所以,从清除血浆中该物质的能力业看,还应该将血浆中浓度(P)这一因素考虑过去。这样,甲肾将该物质从血浆中清除掉的能力为:
U×V/P=[(100mg/100ml)×1ml/min ]/(1mg/100ml)=100ml/min
乙肾为:U×V/P=[(50mg/100ml)×0.8ml/min ]/(0.32mg/100ml)=125ml/min
由此可见,从肾清除血浆中某物质的功能角度来说,乙肾的功能要比甲肾好。
清除率(clearance,C)是指肾在单位时间(一般用每分钟,min)内能将多少毫升血浆中所含的某些物质完全清除出去,这个被完全清除了某物质的血浆毫升数就长为该物质的清除率(ml/min)。其具体计算需要测量三个数值:U(尿中某物质的mg/100ml),V(每分钟尿量,ml/min),P(血浆中某物质的浓度,mg/100ml)。因为尿中该物质均来自血浆,所以,U×V=P×C 亦即 C=U×V/P
根据上式就可计算出各种物质的清除率。例如,Na+清除率的计算方法如下:测得尿量V为1ml/min,尿Na+浓度U为28mmmol/L,血浆Na+浓度P为140mmol/L,Na+清除率
C=[280mmol/L×1ml/min ]/(140mmol/L)=2ml/min
表示肾每分钟清除了2ml血浆中所含的所有Na+。各种物质的清除率并不一样。例如,葡萄糖的清除率为0,因为尿中不含葡萄糖(U=0mg/100ml);而尿素则为70ml/min,等等。因此,清除率能够反映肾对不同物质的清除能力。通过它也可了解肾对各种物质的排泄功能,所以它是一个较好的肾功能测量方法。
这里需要指出,所谓每分钟被完全清除了某物质的血浆毫升数,仅是一个推算的数值,实际上,肾并不一定把某1ml血浆中的某物质完全清除掉,而可能仅仅清除其中的一部分。但是,肾清除该物质的量可以相当于多少毫升血浆中所含的该物质的量。所以说,清除率所表示的血浆毫升数是一个相当量。
二、测定清除率的理论意义
测定清除率不仅可以了解肾的功能,还可能测定肾小球滤过率、肾血流量和推测肾小管转运功能。
(一)测定肾小球滤过率
肾小球滤守率可通过测定菊粉清除率和内生肌酐清除率等方法来测定。
1.菊粉清除率 肾每分钟排出某物质的量(U×V)应为涌小球滤过量与肾小管、集合管的重吸收量和分泌量的代数和。设肾小球滤过率为F;肾小囊囊腔超滤液中某物质(能自由滤过的物质)的浓度(应与血浆中的浓度一致)为P;重吸收量为R;分泌量为E。则U×V=F×P-R+E。如果某物质可以自由滤过,而且既不被重吸收(R=0)也不被分泌(E=0),则U×V=F×P,就可算出肾小球滤过率F。菊粉(inulin,也称菊糖)是符合这个条件的物质,所以它的清除率就是肾小球滤过率。
∵ U×V=F×P
∴ F=U×V/P=C
前文已提出,肾小球滤过率约为125ml/min。这个数值就是根据菊粉的清除率测得的。例如,静脉滴注一定量的菊粉以保持血浆浓度恒定,然后分别测得每分钟尿量(V)为1ml/min,尿中菊粉浓度(U)为125mg/100ml,血浆中菊粉浓度(P)为1mg/100ml,菊粉清除率可用下式计算:
C=U×V/P=[(1ml/min)×(125mg/100ml)]/(1mg/100ml)=125ml/min
所以,肾小球滤过率为125ml/min。
2. 内生肌酐清除率 由于菊粉清除率试验操作复杂,临床上改用较为简便的内生肌酐清除率试验,也以较准确地测得肾小球滤过率。所谓骨生肌酐,是指体内组织代谢所产生的肌酐。试验前二、三日,被试者禁食肉类,以免从食物中摄入过多的外来肌酐。其它饮食照常,但要避免强烈运动或体力劳动,而只从事一般工作。在这种情况下,受试者血浆中的肌酐浓度(平均在1mg/L左右)以及在一昼夜内肌酐的尿中排出总量都比较稳定。这样,在进行肌酐清除率试验时,就不必另给肌酐溶液,只需从第三天清晨起收集24小时的尿,合并起来计算其尿量,并测定混合尿中的肌酐浓度。抽到少量静脉血,测定血浆中的肌酐浓度,按下式可算出24小时的肌酐清除率。
肌酐清除率=[尿肌酐浓度(mg/L)×24小时尿量(L)/血浆肌酐浓度(mg/L)](L/24h)
肌酐能自由通过肾小球滤过,在肾小管中很少被重吸收,但有少量是由近曲小管分泌的。给正常人滴注肌酐,使血浆中浓度高达10-100mg/100ml时,近曲小管分泌肌酐的量增多,此时肌酐清除率可大于菊粉清除率,达175mg/ml。内生肌酐在血浆中的浓度相当低(仅0.1mg/100ml),近曲小管分泌的肌酐量可忽略不计,因此内生肌酐清除率与菊粉清除率相近,可以代表肾小球滤过率。然而,由于测定方法(用苦味酸显色)上的问题,实际测得的数据一般偏低。我国成人内生肌酐清除率平均为128L/24h。
(二)测定肾血流量
如果血浆中某一物质,在经过肾循环一周后可以被完全清除(通过滤过和分泌),亦即在肾动脉中该物质有一定浓度,但在肾静脉中其浓度接近于0,则该物质每分钟的尿中排出量(U×V),应等于每分钟通过肾的血浆中所含的量。设每分钟通过肾的血浆量为X,血浆中该物质浓度为P,即U×V=X×P,则该物质的清除率即为每分钟通过肾的血浆量。
U×V=X×P
C=U×V/P=X
如果在静脉滴注碘锐特(diodrast)或对氨基马尿酸(PAH)的钠盐后维持血浆浓度较低时(1-3mg/100ml),那末当它流经肾时,一次就能被肾几乎全部清除掉,因此,肾静脉中的浓度将接近于0(实际不是0,因为有部分血流通过肾的非泌尿部分)。因此,用此两种物质测得的清除率平均为660ml/min,这一数值代表了肾血浆流量。前述滤过分数就是根据肾小球滤过率和肾血浆流量来推算的。例如
滤过分数=125/660×100≌19%
如果血浆量占全血量的55%,则肾血流量=660/55×100=1200ml/min,约占心输出量的1/5-1/4。
供应肾的血液量应包括供应肾的泌尿部分和非泌尿部分(如肾被膜、肾盂等),而上述测得的肾血浆流量仅代表供应泌尿部分的数值,因此应称为肾有效血浆流量和肾有效血流量。
(三)推测肾小管的功能
通过肾小管滤过率的测定,以及其他物质清除率的测定,可以推测出哪些物质能被肾小管重吸收,哪些物质能被肾小管分泌。
例如,可以自由通过滤过膜的物质,如尿素和葡萄糖,它们的清除率均小于125ml/min(肾小球滤过率),尿素为70ml/min,而葡萄糖为0。这必定是该物质滤过之后遭到了重吸收,其清除率才能小于125ml/min。但是,不能由此而推断说该物质不会被分泌,因为只要重吸收量大于分泌量,其清除率仍可小于125ml/min。
一种物质清除率大于125ml/min(如肌酐的清除率可达175ml/min),这表明这时肾小管必定能分泌该物质,否则基清除率决不可能大于肾小球滤过率。但是,不能由此推断说该物质不会被重吸收,因为只要分泌量大于重吸收量,其清除率仍可大于125ml/min。
(四)自由水清除率
自由水清除率(free water clearance,CH2o),是指单位时间内必须从尿中除去或加入多少容积的纯水(即无溶质的水或称自由水)才能使尿液与血浆等渗,它是定量肾排水能力的指标。例如在水利尿进,血浆渗透浓度(Posm)下降,肾排出大量的低渗尿,尿液渗透浓度(Uosm)小于血奖学金渗透浓度,此时自由水清除率就表示血浆中有一定量的纯水被肾排到等渗尿中,才使尿液稀释和血浆渗透浓度回升。当缺水时,血浆渗透浓度升高,肾排出量少的高渗尿,尿液的渗透浓度大于血浆渗透浓度,此时自由水清除率就表示肾小排出一定量的纯水。这部分纯水保留在血浆,才使尿液浓缩和血浆渗透浓度回降。值得指出的是,血浆中并无真正的自由水存在,自由水清除率是计算出来的。
自由水清除率CH2O=V-Cosm (1)
V为每分钟尿量,Cosm为渗透物质清除率,按下式计算
Cosm=Uosm×V/Posm(2),代入(1)式得
CH2O=V-Uosm×V/Posm=V(1-Uosm/Posm)(3)
由(3)式可见,在等渗尿时,Uosm=Posm,CH2O=0,表示无自由水清除。当在缺水时,Uosm为1100mOsm/kgH2O,尿量V为0.5ml/min,血浆渗透浓度Posm称升高为308mOsm/kgH2O,那么Cosm=1100×0.5/308=1.79ml/min。CH2O=0.5-1.79=-1.29ml/min。自由水清除率为负值,表示肾排出的是浓缩尿、且量少,意味着肾从等渗尿中除去1.29ml/min纯水加入到血浆中,才使尿液浓缩和血浆渗透浓度下降。当水利尿时,尿渗透浓度Uosm为27mOsm/kgH2O,尿量V为20ml/min,血浆渗透浓度Posm为298mOsm/kgH2O,那么Cosm=27×20/298=1.81ml/min,CH2O=20-1.81=18.1ml/min,自由水清除率为正值,表示肾排出大量稀释尿,意味着肾从血浆中除去18.1ml/min纯水加入到等渗尿液中,才使尿液稀释和血浆渗透浓度升高。
第七节 尿的排放
尿的生成是个连续不断的过程。持续不断进入肾盂的尿液,由于压力差以及肾盂的收缩而被送入输尿管。输尿管中的尿液则通过输尿管的周期性蠕动而被送入到膀胱。但是,膀胱的排尿(micturition)是间歇地进行的。尿液在膀胱内贮存并达到一定量时,才能引起反射性排尿动作,将尿液经尿道排放于体外。
一、膀胱与尿道的神经支配
膀胱逼尿肌和内括约肌受交感神经和副交感神经支配。由2-4骶髓发出的盆神经中含副交感神经纤维,它的兴奋可使逼尿肌收缩、膀胱内括约肌松弛,促进排尿。交感神经纤维是由腰髓发出,经腹下神经到达膀胱。它的兴奋则使逼尿肌松弛、内括约肌收缩,阻抑尿的排放。但在排尿活动中交感神经的作用比较次要。
膀胱外括约肌受阴部神经(由骶髓发出的躯体神经)支配,它的兴奋可使外括约肌收缩。这一作用受意识控制。至于外括约肌的松弛,则是阴部神经活动的反射性抑制所造成的。
上述三种神经也含有传入纤维。膀胱充胀感觉的传入纤维在盆神经中;传导膀胱痛觉的纤维在腹下神经中;而传导尿道感觉的传入纤维在阴部神经中(图8-22)。
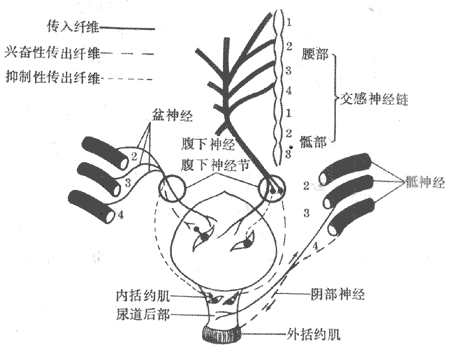
图8-22 膀胱和尿道的神经支配
二、排尿反射
在正常情况下,膀胱逼尿肌在副交感神经紧张冲动的影响下,处于轻度收缩状态,使膀胱内压经常保持在0.98kPa(10cm H2O),因为膀胱具有较大的伸展性,导致内压稍升高后可以很快回降。当尿量增加到400-500ml时膀胱内压才超过0.98kPa(10cm H2O)而明显升高(图8-23)。如果膀胱内尿量增加到700ml,膀胱内压随之增加至3.43kPa(35cm H2O)时,逼尿肌便出现节律性收缩,排尿欲也明显增加,但此时还可有意识地控制排尿。当膀胱内压达到6.86kPa(70cm H2O)以上时,便出现明显的痛感以致不得不排尿。
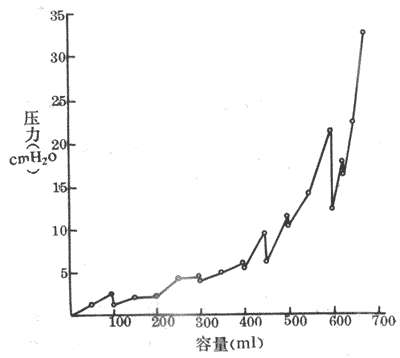
图8-23 人膀胱充盈过程中膀胱容量与压力的关系
图中压力垂直降低,表示容量恒定时膀胱的适应程度
排尿活动是一种反射活动。当膀胱尿量充盈到一定程度时(400-500ml),膀胱壁的牵张感受器受到刺激而兴奋。冲动沿盆神经传入,到达骶髓的排尿反射初级中枢;同时,冲动也到过脑干和大脑皮层的排尿反射高位中枢,并产生排尿欲。排尿反射进行时,冲动沿盆神经传出,引起逼尿肌收缩、内括约肌松弛,于是尿液进入后尿道。这时尿液还可以刺激尿道的感受器,冲动沿阴部神经再次传到脊髓排尿中枢,进一步加强其活动,使外括约肌开放,于是尿注被强大的膀胱内压(可高达14.7kPa(150cm H2O)驱出。尿液对尿道的刺激可进一步反射性地加强排尿中枢活动。这是一种正反馈,它使排尿反射一再加强,直至尿液排完为止。在排尿末期,由于尿道海绵体肌肉收缩,可将残留于尿道的尿液排出体外。此外,在排尿时,腹肌和膈肌的强大收缩也产生较高的腹内压,协助克服排尿的阻力。
大脑皮层等排尿反射高位中枢能对脊髓初级中枢施加易化或抑制性影响,以控制排尿反射活动。小儿大脑发育未臻完善,对初级中枢的控制能力较弱,所以小儿排尿次数多,且易发生在夜间遗尿现象。
排尿或贮尿任何一方发生障碍,均可出现排尿异常,临床上常见的有尿频,尿潴留和尿失禁。排放次数过多者称为尿频,常常是由于膀胱炎症或机械性刺激(如膀胱结石)而引起的。膀胱中尿液充盈过多而不能排出者称为尿潴留。尿潴留多半是由于腰骶部脊髓损伤使排尿反射初级中枢的活动发生障碍所致。但尿流受阻也能造成尿潴留。当脊髓受损,以致初级中枢与大脑皮层推动功能联系时,排尿便失去了意识控制,可出现尿失禁。
参考资料
1.徐丰彦,张镜如主编.人体生理学,北京:人民卫生出版社,1989
2.何小瑞,姚泰.管球反馈对肾小球血流动力学的影响及其机制.生理学进展 1991;22:216-220
3.潘敬运.肾神经的功能.生理科学进展 1985;16:215-219
4.Berne RM,Levy MN. Physiology 3rded, pp 719-753,CV Mosby Co,St Louis,1993
5.Stokes -J-B.Electroneutral NaCItransport in the distal tubule.Kidney International 1989;36:427
6.Rose BD.Diuretics.KidneyInternational 1991;39:336
7.Doucet A.Na-K-ATPase in thekidney tubule in relation to natriuresis.Kidney International 1992;41:Sll8
8.Sands JM,KokkoJP.Countercurrent system.Kidney International 1990;38:695
9.Molony DA,Reeves WB,AndreoliTE.Na+: K+;2CI-cotransport and thick ascendinglimb.kidney International 1989;36:418
10.Stokes J-B.Sodium and potassiumtransport by the collecting duct.Kidney International 1990;38:678

 中医世家
中医世家 浦 标 网
浦 标 网 河南大学精品课程
河南大学精品课程 图书资料室
图书资料室