神经组织(nerve tissue)构成神经系统。神经系统分中枢神经系统(脑与脊髓)和周围神经系统(神经和神经节)两大部分,两者是相互联系的整体。神经组织是由神经细胞(nerve cell)和神经胶质细胞(neuroglial cell)组成的,它们都是有突起的细胞。神经细胞是神经系统的结构和功能单位,亦称神经元。神经元数量庞大,整个神经系统约有1011个,它们具有接受刺激、传导冲动和整合信息的能力。神经元的突起以特化的连接结构――突触彼此连接,形成复杂的神经通路和网络,将化学信号或电信号从一个神经元传给另一个神经元,或传给其组织的细胞,使神经系统产生感觉和调节其他系统的活动,以适应内、外环境的瞬息变化。有些神经元还有内分泌功能。神经胶质细胞的数量比神经元更多,但不具有神经元的上述特性,它们的功能是对神经元起支持、保护、分隔、营养等作用,两者的关系十分密切。
神经元的胞体主要分布在中枢神经系统,如大脑皮质、小脑皮质、脑内众多的神经核团和脊髓灰质;也存在于周围神经系统的神经节内,如脑神经节、脊神经节、植物神经节。神经元的突起则组成中枢神经系统的神经通路和神经网络以及遍布在的神经。分布到体表和骨骼肌的神经躯体神经(somatic nerve)分布到内脏、心血管和腺体的称内脏神经(visceral nerve)或植物神经(vegetative nerve);植物神经又分交感神经和副交感神经,分别与相应的植物神经节相连。
一、神经元
神经元(neuron)的形态多种多样,但都可分为胞体(soma)和突起(neurite)两部分(图7-1)。胞体的大小差异很大,小的直径仅5~6μm,大的可达100μm以上,突起的形态、数量和长短也很不相同。神经元突起又分树突(dendrite)和轴突(axon)两种。树突多呈树状分支,它可接受刺激并将冲动传向胞体;轴突呈细索状,末端常有分支,称轴突终末(axon terminal),轴突将冲动从胞体传向终末。通常一个神经元有一个至多个树突,但轴突只有一条(图7-2)。神经元的胞体越大,其轴突越长。
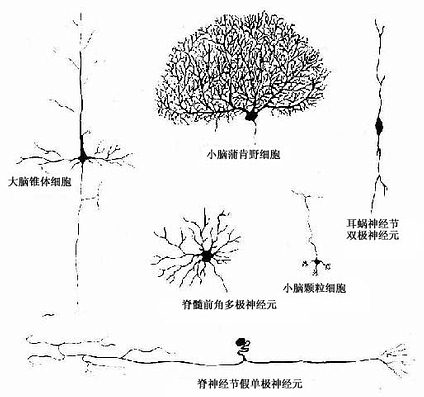
图7-1 神经元的几种主要形态类型
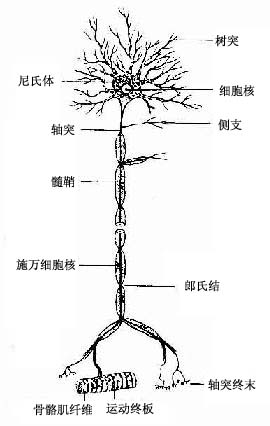
图7-2 运动神经元模式图
(一)神经元的分类
神经元有几种分类法。根据突起的多少可将神经元分为三种:①多极神经元(multipolar neuron),有一个轴突和多个树突;②双极神经元(bipolar neuron),有两个突起,一个是树突,另一个是轴突;③假单极神经元(pseudounipolar neuron),从胞体发出一个突起,距胞体不远又呈“T”形分为两支,一支分布到外周的其他组织的器官,称周围突(peripheral process);另一支进入中枢神经系统,称中枢突(central process)(图7-3)。假单极神经元的这两个分支,按神经冲动的传导方向,中枢突是轴突,周围突是树突;但周围突细而长,与轴突的形态类似,故往往通称轴突。
根据轴突的长短,神经元可分为:①长轴突的大神经元,称GolgiⅠ型神经元,最长的轴突达1m以上;②短轴突的小神经元,称GolgiⅡ型神经元,轴突短的仅数微米。
根据神经元的功能又可分:①感觉神经元(sensory neuron),或称传入神经元(afferent neuron)多为假单极神经元,胞体主要位于脑脊神经节内,其周围突的末梢分布在皮肤和肌肉等处,接受刺激,将刺激传向中枢。②运动神经元(motor neuron),或称传出神经元(efferent neuron)多为多极神经元,胞体主要位于脑、脊髓和植物神经节内,它把神经冲动传给肌肉或腺体,产生效应。③中间神经元(interneuron),介于前两种神经元之间,多为多极神经元(图7-3)。动物越进化,中间神经元越多,人神经系统中的中间神经元约占神经元总数的99%,构成中枢神经系统内的复杂网络。
根据神经元释放的神经递质(neurotransmitter),或神经调质(neuromodulator),还可分为:①胆碱能神经元(cholinergic neuron);②胺能神经元(aminergic neuron);③肽能神经元(peptidergic neuron);④氨基酸能神经元。
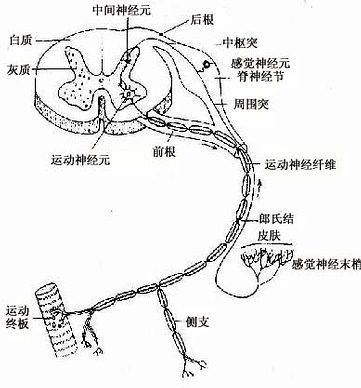
图7-3 脊髓及脊神经,示三种神经元的关系
(二)神经元的结构
1.细胞膜 神经元的细胞膜是可兴奋膜(excitable membrane),它在接受刺激、传播神经冲动和信息处理中起重要作用。通常是神经元的树突膜和胞体膜接受刺激或信息,轴突膜(轴膜)传导神经冲动。神经元细胞膜的性质决定于膜蛋白的种类、数量、结构和功能,其中有些膜蛋白是离子通道(ionic channel)、按所通过的离子分别命名为钠信道、钾信道或钙信道等;还有一些膜蛋白是受体(receptor),可与相应的化学物质(神经递质)结合,使离子通道开放。目前认为,控制离子通道的开闭存在一种闸门机制(gating mechanism),有些通道是受电刺激而开放的,称电位门控通道(voltage-gated channel),有些是当化学物质与受体结合时才开放的,称化学门控通道(chemically-gated channel)。还有一些通道不受上述机制控制,而是经常开放着的。一般是轴膜富含电位门控通道,树突膜和胞体膜主要是化学门控通道。
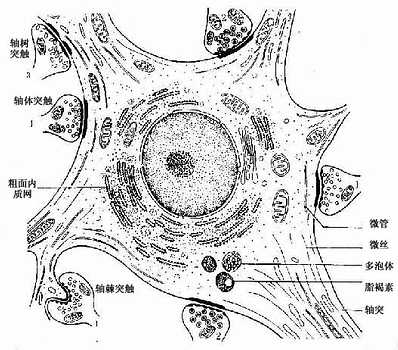
图7-4 多极神经元及其突触超微结构模式图
1.突触扣结内有圆形清亮小泡,内含乙酰胆碱
2、突触扣结内有颗粒型小泡,内含单胺类
3、突触扣结内有扁平清亮小泡,内含甘氨酸等
2.胞体 神经元胞体是细胞的营养中心。胞体的中央有一个大而圆的细胞核,核异染色质少,故着色浅,核仁大而明显。胞体的细胞质称核周质(perikaryon),含有较发达的粗面内质网、游离核糖体、微丝、神经丝和微管以及高尔基复合体等(图7-4)。粗面内质网常呈现规则的平行排列,游离核糖体分布于其间,它们在光镜下呈嗜碱性颗粒或小块,称尼氏体(Nissl bodies)。大神经元尤其是运动神经元的尼氏体丰富而粗大,呈斑块状(图7-5);小神经元的尼氏体则较小而少。大神经元胞体内含大量尼氏体和发达的高尔基复合体,表明细胞具有合成蛋白质的旺盛功能。合成的蛋白质包括复制细胞器所需蛋白质和产生神经递质有关的酶等。神经丝(neurofilament)直径约为10nm,是中间丝的一种,常集合成束,微管直径约25nm,它常与神经丝交叉排列成网,并伸入树突和轴突内,构成神经元的细胞骨架(cytoskeleton),参与物质运输。在银染色切片中,神经丝和微管呈棕黑色细丝,又称神经原纤维(neurofibril)(图7-6)。胞体内还含有色素,最常见的是棕黄色的脂褐素(lipofuscin),并随年龄而增多。
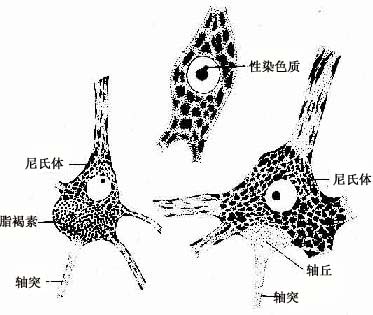
图7-5 神经元胞体光镜结构,示尼氏体
图7-6 神经元银染色,示神经原纤维
某些神经元,如下丘脑具内分泌功能的分泌神经元(secretory neuron),胞体内含直径100~300nm的分泌颗粒,颗粒内含肽类激素(如加压素、催产素等)。
3.树突 树突内的结构与核周质基本相似。在树突分支上常见许多棘状的小突起,称树突棘(dendritic spine)。树突棘是神经元之间形成突触的主要部位,电镜下可见树突棘内有2~3层滑面内质网形成的板层,板层间有少量致密物质,称此为棘器(spine apparatus)(图7-7)。树突棘的数量及分布因不同神经元而异,并可随功能而改变。在大脑皮质锥体细胞和小脑皮质蒲肯野细胞的树突上,树突棘数量最多而明显,一个蒲肯野细胞的树突棘可多达10万个以上。树突的功能主要是接受刺激,树突棘和树突使神经元的接受面大为扩大。
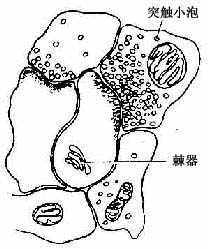
图7-7 树突棘及棘器模式图
4.轴突 轴突通常自胞体发出,但也有从主树突干的基部发出。胞体发出轴突的部位常呈圆锥形,称轴丘(axon hillock),光镜下此区无尼氏体,染色淡(图7-5)。轴突的长短不一,短者仅数微米,长者可达一米以上。轴突一般比树突细,全长直径较均一,有侧支呈直角分出。轴突表面的细胞膜称轴膜(axolemma),内含的胞质称轴质(axoplasm)。轴质内有大量微管和神经丝,此外还有微丝、线粒体、滑面内质网和一些小泡等。微管与神经丝均很长,沿轴突长轴平行排列。微丝较短,主要分布于轴膜下,常与轴膜相连。电镜观察轴突冷冻蚀刻标本,可见微丝、微管和神经丝之间均有横桥连接,构成轴质中的网架结构。轴突内无尼氏体和高尔基复合体,故不能合成蛋白质,轴突成分的更新及神经递质全成所需的蛋白质和酶,是在胞体内合成后输送到轴突及其终末的。
轴突的主要功能是传导神经冲动。神经冲动的传导是在轴膜上进行的,轴突起始段轴膜的电兴奋性阈较胞体或树突低得多,故此处常是神经元发生冲动的起始部位。轴突起始段长约15~25μm,电镜下见轴膜较厚,膜下有电子密度致密层。
轴突内的物质运输称轴突输送(axonal transport)。神经元胞体内新合成的微管、微丝和神经丝组成的网架缓慢地移向轴突终末(0.1~0.4mm/天),称此为慢速输送。另外还有一种快速双向的轴突输送(100~400mm/天)。轴膜更新所需的蛋白质、含神经递质的小泡及合成递质所需的酶等,由胞体输向终末,称快速顺向轴突输送。轴突终末代谢产物或由轴突终末摄取的物质(蛋白质、小分子物质或由邻细胞产生的神经营养因子等)逆行输向胞体,称快速逆向轴突输送(图7-8)。某些微生物或毒素(如破伤风毒素、狂犬病毒)进入轴突终末,也可通过逆行性运输迅速侵犯神经元胞体,新近的研究表明,微管在轴突输送中起重要作用,微管与轴质中的动力蛋白(dynein)或激蛋白(kinesin)相互作用,可推动小泡向一定方向移动。此外微丝也与轴突输送作用有关。
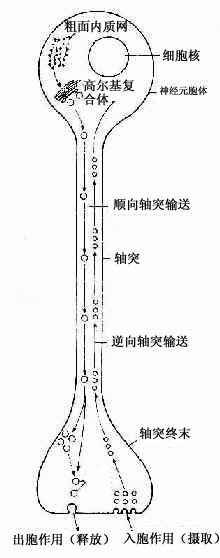
图7-8 双向轴突输送示意图
二、突触
突触(synapse)是神经元传递登记处的重要结构,它是神经元与神经元之间,或神经元与非神经细胞之间的一种特化的细胞连接,通过它的传递作用实现细胞与细胞之间的通讯。在神经元之间的连接中,最常见是一个神经元的轴突终末与另一个神经元的树突、树突棘或胞体连接,分别构成轴-树(axodendritic)、轴-棘(axospinous)、轴-体(axosomatic)突触(图7-4)。此外还有轴-轴(axoaxonal)和树-树(dendrodendritic)突触等。突触可分为化学突触(chemical synapse)和电突触(electrical synapse)两大类。前者是以化学物质(神经递质)作为通讯的媒介,后者是亦即缝隙连接,是以电流(电讯号)传递信息。哺乳动特神经系统以化学突触占大多数,通常所说的突触是指化学突触而言。
突触的结构可分突触前成分(presynaptic element)、突触间隙(synaptic cleft)和突触后成分(postsynaptic element)三部分。突触前、后成分彼此相对的细胞膜分别称为突触前膜和突触后膜(presynaptic and postsynapticmembrane),两者之间在宽约15~30nm的狭窄间隙为突触间隙,内含糖蛋白和一些细丝。突触前成分通常是神经元的轴突终末,呈球状膨大,它们在银染色标本中呈现为棕黑色的环扣状,附着在另一神经元的胞体或树突上,称突触扣结(synaptic bouton)(图7-9)。
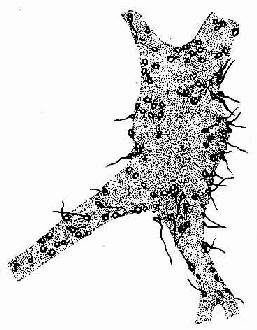
图7-9 脊髓运动神经元银染,示突触扣结
电镜下,突触扣结内含许多突触小泡(synapse vesicle),还有少量线粒体、滑面内质网、微管和微丝等(图7-10,7-11)。突触小泡的大小和形状不一,多为圆形,直径40~60nm,亦有的呈扁平形。突触小泡有的清亮,有的含有致密核芯(颗粒型小泡),大的颗粒型小泡直径可达200nm(图7-4)。突触小泡内含神经递质或神经调质。突触前膜和后膜均比一般细胞膜略厚,这是由于其胞质面附有一些致密物质所致(图7-10)。在突触前膜还有电子密度高的锥形致密突起(dense projection)突入胞质内,突起间容纳突触小泡。突触小泡表面附有突触小泡相关蛋白,称突触素Ⅰ(synapsinⅠ),它使突触小泡集合并附在细胞骨架上。突触前膜上富含电位门控通道,突触后膜上则富含受体及化学门控通道。当神经冲动沿轴膜传至轴突终末时,即触发突触前膜上的电位门控钙通道开放,细胞外的Ca2+进入突触前成分,在ATP的参与下使突触素I发生磷酸化,促使突触小泡移附在突触前膜上,通过出胞作用释放小泡内的神经递质到突触间隙内。其中部分神经递质与突触后膜上相应受体结合,引起与受体偶联的化学门控通道开放,使相应离子进出,从而改变突触后膜两侧离子的分布状况,出现兴奋或抑制性变化,进而影响突触后神经元(或非神经细胞)的活动。使突触后膜发生兴奋的突触称兴奋性突触(excitatory synapse),使突触后膜发生抑制的称抑制性突触(inhibitory synapse)。突触的兴奋或抑制,取决于神经递质及其受体的种类。
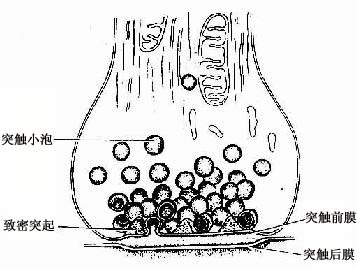
图7-10 化学突触超微结构模式图
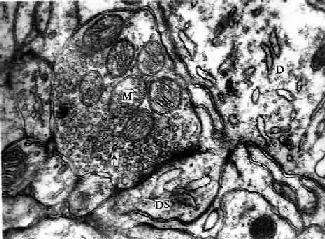
图7-11 豚鼠 小脑电镜像示突触
D树突,DS树突棘,M线粒体,↑突触小泡
(白求恩医科大学尹昕、朱秀雄供图)
一个神经元既可与其他神经元建立许多突触连接,亦可接受来自其他神经元的许多突触信息。一个神经元上突触数目的多少视不同的神经元而有很大差别,例如小脑的颗粒细胞只有几个突触,一个运动神经元要有1万个左右突触,而小脑的蒲肯野细胞树突上的突触就有10万个以上。一个神经元上众多的突触中,有些是兴奋性的,有些则是抑制性的。如果所有兴奋性突触活动的总和超过抑制性突触活动的总和,并足以刺激该神经元的轴突起始段产生动作电位时,则该神经元发生兴奋;反之,则表现为抑制。
根据突触前、后膜胞质面致密物质厚度的差异,可把突触分为Ⅰ、Ⅱ两型。Ⅰ型突触的突触后膜胞质面附有的致密物质较突触前膜的厚,两者不对称,突触间隙也较宽(30nm),故称不对称突触。Ⅱ型突触前、后膜的致密物质少,厚度相近,突触间隙较窄(20nm),称对称突触。有人认为Ⅰ型属兴奋性突触,Ⅱ型属抑制性突触。
神经递质的种类很多,包括:乙酰胆碱(acetylcholine,Ach);单胺类,如去甲肾上腺素(norepinephrine),多巴胺(dopamine,DA)和5-羟色胺(5-hydroxytryptamine,5-HT);氨基酸类,如γ-氨基丁酸(γ-aminobutyric acid GABA)、甘氨酸(glycine)、谷氨酸(glutamic acid)等。新近又发现大量的神经肽(neuropeptide),如P物质(substance P)、脑啡肽(enkephalin)、神经隆压素(neurotensin)、血管活性肠肽(vasoactive intestinal polypeptide,VIP)、胆囊收缩素(cholesystokinin)、加压素(vasopressin)、和下丘脑释放激素(hypothalamic releasing hormones)等约40多种。有些神经肽亦见于胃肠管的内分泌细胞,故总称为脑肠肽(braingut peptide)。这些肽类物质能改变神经元对经典神经递质的反应,起修饰经典神经递质的作用,故称为神经调质(neuromodulator)。不同形态大小的突触小泡所含的神经递质也不同,如圆形清亮小泡多数含乙酰胆碱,小颗粒型小泡含单胺类,大颗粒型小泡往往含神经肽。过去认为一个神经元一般只产生和释放一种神经递质,但近来应用免疫细胞化学法研究发现,某些神经元可产生和释放两种或两种以上的神经递质和(或)神经调质,其中一种往往是经典神经递质(Ach或NE),另一种则是神经肽。神经递质或神经调质共存的生理意义尚待研究。
突触后膜上的受体是一种膜蛋白,它能与相应的神经递质的结合而使突触后膜产生兴奋或抑制。神经递质的种类很多,受体的种类相应也很多。虽然一种受体只与相应的一种神经递质结合,但一种神经递质却可有不止一种受体。如乙酰胆碱受体就有N型(兴奋型)和M型(多数为兴奋型,少数为抑制型),去甲肾上腺素受体亦有α和β两类。所以,突触的兴奋或抑制,不仅取决于神经递质的种类(如γ-氨基丁酸是脑内一种抑制性神经递质),更重要的还取决于受体的类型。同一种神经递质在神经系统的不同部位有兴奋或抑制的不同效应,主要原因是突触后膜上受体类型的不同。突触后膜上的受体可直接与通道蛋白偶联或其本身就是一种通道蛋白,故当神经递质与这种形式的受体结合后使突触后膜产生的兴奋或抑制性变化十分迅速,所以把这种形式的受体称为快速作用受体,如乙酰胆碱N型受体属于快速作用受体。另外亦有慢速作用受体,这种受体一般与G蛋白偶联,再经过细胞内第二信使(环腺苷酸,cAMP)及蛋白磷酸化产生效应,它的作用比前者缓慢,但能把递质-受体相互作用所提供的微弱信号放大数千倍,如去甲肾上腺素β受体属于这种慢速作用受体。
释放到突触间隙的递质分子与突触后膜的受体结合产生生理效应后,很快便被相应的酶灭活(如Ach),或吸收入突触终末内被分解(如NE),以迅速消除该递质的作用,这样才能保证突触传递的灵敏性。递质的分解产物可被重新利用合成新的递质。非肽类递质除在胞体合成外,通常也可在轴突终末内合成,而肽类递质则只能在胞体内合成,释放后亦不能回收。合成的递质分子一般都贮存在突触小泡内。
三、神经胶质细胞
神经胶质细胞或简称胶质细胞(glial cell),广泛分布于中枢和周围神经系统,其数量比神经元的数量大得多,胶质细胞与神经元数目之比约10:1~50:1。胶质细胞与神经元一样具有突起,但其胞突不分树突和轴突,亦没有传导神经冲动的功能。胶质细胞可分几种,各有不同的形态特点,但HE染色只能显示其细胞核,用特殊的金属浸镀技术(银染色)或免疫细胞化学方法可显示细胞的全貌(图7-12)。
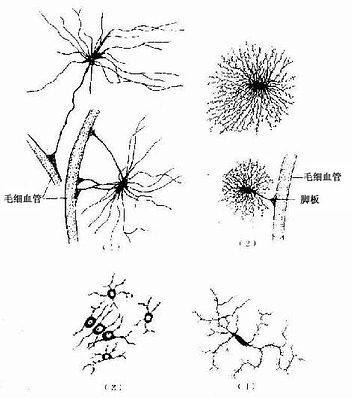
图7-12 中枢神经的几种胶质细胞(银染法)
(1)纤维性星形胶质细胞(2)原浆性星形胶质细胞
(3)少突胶质细胞(4)小胶质细胞
(一)中枢神经系统的胶质细胞
1.星形胶质细胞 星形胶质细胞(astrocyte)是胶质细胞中体积最大的一种,与少突胶质细胞合称为大胶质细胞(macroglia)。细胞呈星形,核圆形或卵圆形,较大,染色较浅(图7-13)。星形胶质细胞可分两种:①纤维性星形胶质细胞(fibrous astrocyte),多分布在白质,细胞的突起细长,分支较少,胞质内含大量胶质丝(glial fiament)。组成胶质丝的蛋白质称胶质原纤维酸性蛋白(glial fibrillary acidicprotein,GFAP),用免疫细胞化学染色技术,能特异性地显示这类细胞。②原浆性星形胶质细胞(protoplasmic astrocyte),多分布在灰质,细胞的突起较短粗,分支较多,胞质内胶质丝较少。星形胶质细胞的突起伸展充填在神经元胞体及其突起之间,起支持和分神经元的作用。有些突起末端形成脚板(end feet),附在毛细血管壁上(图7-12),或附着在脑和脊髓表面形成胶质界膜(glia limitans)(图7-14)。
星形胶质细胞之间的细胞间隙狭窄而迂回曲折,宽约15~20nm,内含组织液,神经元借此进行物质交换(图7-14),星形胶质细胞能吸收细胞间隙的K+,以维持神经元周围环境K+含量的稳定性,它还能摄取和代谢某些神经递质(如γ-氨基丁酸等),调节细胞间隙中神经递质的浓度,有利神经元的活动。在神经系统发育时期,某些星形胶质细胞具有引导神经元迁移的作用,使神经元到达预定区域并与其他细胞建立突触连接。中枢神经系统损伤时,星形胶质细胞增生、肥大、充填缺损的空隙,形成胶质瘢痕(glial scar)。
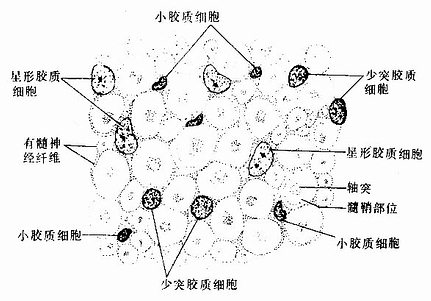
图7-13 中枢神经系各种神经胶质细胞的细胞核及神经纤维横切
(脊髓白质,Nissl法染色)
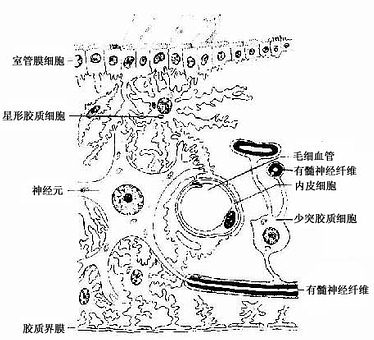
图7-14 中枢神经胶质细胞与神经元和毛细胞血管的关系图解
2.少突胶质细胞 在银染色标本中,少突胶质细胞(oligodendrocyte)的突起较少(图7-12),但用特异性的免疫细胞化学染色,是可见少突胶质细胞的突起并不很少,而且分支也多。少突胶质细胞的胞体较星形胶质细胞的小,核圆,染色较深(图7-13)。胞质内胶质丝很少,但有较多微管和其他细胞器。少突胶质细胞分布在神经元胞体附近和神经纤维周围,它的突起末端扩展成扁平薄膜,包卷神经元的轴突形成髓鞘,所以它是中枢神经系统的髓鞘形成细胞(图7-18)。新近研究认为,少突胶质细胞还有抑制再生神经元突起生长的作用。
3.小胶质细胞 小胶质细胞( microglia)是胶质细胞中最小的一种。胞体细长或椭圆,核小,扁平或三角形,染色深(图7-13)。细胞的突起细长有分支,表面有许多小棘突(图7-12)。小胶质细胞的数量少,约占全部胶质细胞的5%左右。中枢神经系统损伤时,小胶质细胞可转变为巨噬细胞,吞噬细胞碎屑及退化变性的髓鞘。血循环中的单核细胞亦侵入损伤区,转变为巨噬细胞,参与吞噬活动。由于小胶质细胞有吞噬功能,有人认为它来源于血液中的单核细胞,属单核吞噬细胞系统。
4.室管膜细胞 室管膜细胞(ependymal cell)为立方或柱形,分布在脑室及脊髓中央管的腔面,形成单层上皮,称室管膜(ependyma)。室管膜细胞表面有许多微绒毛,有些细胞表面有纤毛(图7-14)。某些地方的室管膜细胞,其基底面有细长的突起伸向深部,称伸长细胞(tanycyte)。
(二)周围神经系统的胶质细胞
1.施万细胞 施万细胞(Schwann cell)是周围神经纤维的鞘细胞,它们排列成串,一个接一个地包裹着周围神经纤维的轴突。在有髓神经纤维,施万细胞形成髓鞘,是周围神经系统的髓鞘形成细胞。施万细胞外表面有一层基膜,在周围神经再生中起重要作用。
2.卫星细胞 卫星细胞(satellite cell)是神经节内包裹神经元胞体的一层扁平或立方形细胞,故又称被囊细胞。细胞核圆或卵圆形,染色较深。细胞外面有一层基膜。
四、神经纤维和神经
(一)神经纤维
神经纤维(nerve fiber)是由神经元的长轴突外包胶质细胞所组成。包裹中枢神经纤维轴突的胶质细胞是少突胶质细胞,包裹周围神经纤维轴突的是施万细胞。根据包裹轴突的胶质细胞是否形成髓鞘(myelin sheath),神经纤维可分有髓神经纤维(myelinated fiber)和无髓神经纤维(unmyelinated nerve fiber)(图7-15)。神经纤维主要构成中枢神经系统的白质和周围神经系统的脑神经、脊神经和植物神经。
1.有髓神经纤维
(1)周围神经系统的有髓神经纤维:这种神经纤维的轴突,除起始段和终末外均包有髓鞘(图7-2,7-3)。髓鞘分成许多节段,各节段间的缩窄部称郎氏结(Ranvier node)。轴突的侧支均自郎氏结处发出。相邻两个郎氏结之间的一段称结间体(internode)。轴突越粗,其髓鞘也越厚,结间体也越长。每一结间体的髓鞘是由一个施万细胞的胞膜融合,并呈同心圆状包卷轴突而形成的,电镜下呈明暗相间的同心状板层(图7-16,7-17)。髓鞘的化学成分主要是类脂和蛋白质,称髓磷脂(myelin)。髓磷脂中类脂含量很高,约占80%,故新鲜髓鞘呈闪亮的白色,但在常规染色标本上,因类脂被溶解,仅见残留的网状蛋白质(图7-15)。若标本用锇酸固定和染色,髓磷脂保存,髓鞘呈黑色,在其纵切面上常见一些漏斗型的斜裂,称施-兰切迹(Schmidt-Lantermann incisure)(图7-15)。

图7-15 周围神经纤维
(1)神经铺片示有髓和无髓神经纤维
(2)锇酸固定染色示髓鞘与施兰切迹
(3)银染法示郎氏结
施万细胞的胞核呈长卵圆形,其长轴与轴突平行,核周有少量胞质。由于施万细胞包在轴突的外面,故又称神经膜细胞(neurilemmal cell),它的外面包有一层基膜。施万细胞最外面的一层胞膜与基膜一起往往又称神经膜(neurilemma),光镜下可见此膜。
髓鞘的形成:在有髓神经纤维发生中,伴随轴突一起生长的施万细胞表面凹陷成一纵沟,轴突位于纵沟内,沟缘的胞膜相贴形成轴突系膜(mesaxon)。轴突系膜不断伸长并反复包卷轴突,把胞质挤至细胞的内、外边缘及两端(即靠近郎氏结处),从而形成许多同心圆的螺旋膜板层,即为髓鞘(图7-16,7-17)。故髓鞘乃成自施万细胞的胞膜,属施万细胞的一部分。施万细胞的胞质除见于细胞的外、内边缘和两端外,还见于髓鞘板层内的施-兰切迹。该切迹构成螺旋形的胞质通道,并与细胞外、内边缘的胞质相通。

图7-16 周围神经纤维髓鞘形成及鞭超微结构模式图
(1)(2)(3)髓鞘发生过程(4)有髓神经纤维超微结构
(5)无髓神经纤维超微结构

图7-17 人有髓神经纤维髓鞘电镜像×40000(中山医科大学张爱华供图)
(2)中枢神经系统的有髓神经纤维:其结构基本与周围神经系统的有髓神经纤维相同,不同的是它的髓鞘不是施万细胞,而是由少突胶质细胞突起末端的扁平薄膜包卷轴突而形成。一个少突胶质细胞有多个突起可分别包卷多个轴突,其胞体位于神经纤维之间(图7-18)。其次是中枢有髓神经的外表面没有基膜包裹,髓鞘内亦无施-兰切迹。
有髓神经的轴膜兴奋是呈跳跃式传导的,故传导速度快。
有髓神经纤维的髓鞘,因含高浓度类脂而具嫌水性,它不容带离子的水溶液通过而起绝缘作用。有髓神经纤维轴突的轴膜,除轴突起始段和轴突终末外,只有在郎氏结处才暴露于细胞外环境。其余大部分的轴膜均被髓鞘包裹。由于髓鞘的电阻比轴膜高得多,而电容却很低,通过轴突的电流只能使郎氏结处的轴膜去极化而产生兴奋。所以,从轴突起始段产生的神经冲动(动作电位)的传导,是通过郎氏结处的轴膜进行的,即从一个郎氏结跳到下一个郎氏结,呈快速的跳跃式传导。故而,结间体越长,跳跃的距离也越大,传导速度也就越快。
2.无髓神经纤维
(1)周围神经系统的无髓神经纤维:由较细的轴突和包在它外面的施万细胞组成。施万细胞沿着轴突一个接一个地连接成连续的鞘,但不形成髓鞘,故无郎氏结;而且一个施万细胞可包裹许多条轴突(图7-16,7-19)。施万细胞外面亦有基膜。
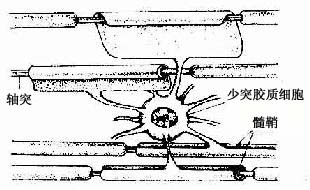
图 7-18 少突胶质细胞与中枢有髓神经纤维关系模式图
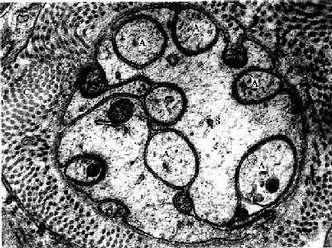
图7-19 大鼠无髓神经纤维横切电镜像×45000
A轴突,S施万细胞,C胶原原纤维
(白求恩医科大学尹昕、朱秀雄教授供图)
(2)中枢神经系统的无髓神经纤维:轴突外面没有任何鞘膜,因此是裸露的轴突,它们与有髓神经纤维混杂在一起。在一些脑区,它们可被星形胶质细胞的突起分隔成束。
无髓神经纤维因无髓鞘和郎区结,电流通过轴膜是沿着轴突连续传导的,故其传导速度比有髓神经纤维慢得多。
(二)神经
周围神经系统的神经纤维集合在起,构成神经(nerve),分布到全身各器官和组织。一条神经内可以只含有感觉(传入)神经纤维或运动(传出)神经纤维,但大多数神经是同时含有感觉、运动和植物神经纤维的。在结构上,多数神经同时含有髓和无髓两种神经纤维。由于有髓神经纤维的髓鞘含髓磷脂,故神经通常呈白色。
包裹在神经外面的致密结缔组织称神经外膜(epineurium)。神经内的神经纤维,又被结缔组织分隔成大小不等的神经纤维束,包裹每束神经纤维的结缔组织称神经束膜(perineurium)(图7-20)。神经束膜的外层是结缔组织,内层则由多层的扁平上皮细胞组成,称神经束膜上皮(perineural epithelium),上皮细胞之间有紧密连接,每层上皮都有基障进入神经内部。神经纤维束内的每条神经纤维又有薄层疏松结缔组织包裹,称神经内膜(endoneurium)。神经内的血管较丰富,神经外膜内的纵行血管发出分支进入神经束膜,进而在神经内膜形成毛细血管网。神经内膜亦含有淋巴管。
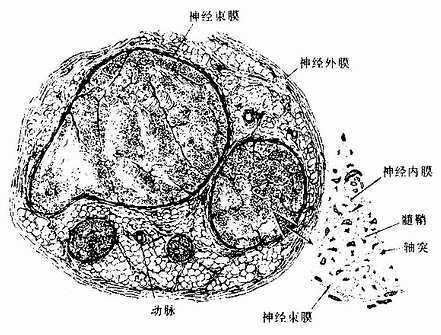
图7-20 坐骨神经横切,示神经外膜、神经束膜与神经内膜
五、神经末梢
周围神经纤维的终末部分终止于全身各种组织或器官内,形成各式各样的神经末梢(nerve ending),按其功能可分感觉神经末梢和运动神经末梢两大类。
(一)感觉神经末梢
感觉神经末梢(sensory nerve ending)是感觉神经元(假单极神经元)周围突的终末部分,该终末与其他结构共同组成感受器。感受器能接受内、外环境的各种刺激,并将刺激转化为神经冲动,传向中枢,产生感觉。感觉神经末梢按其结构可分游离神经末梢和有被囊神经末梢两类。
1.ng)结构较简单。较细的有髓或无髓神经纤维的终末部分失去施万细胞,裸露的轴突末段分成细支,分布在表皮、角膜和毛囊的上皮细胞间,或分布在各型结缔组织内,如骨膜、脑膜、血管外膜、关节囊、肌腱、韧带、筋膜和牙髓等处。此类末梢感受冷、热、轻触和痛的刺激(图7-21,7-3)。
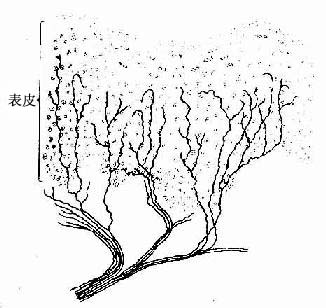
图7-21 表皮内的游离神经末梢
2.有被囊神经末梢 有被囊神经末梢(encapsulated nerve ending)外面均包裹有结缔组织被囊,它们的种类很多,常见的有如下几种:
(1)触觉小体(tactile corpuscle)又称Meissner小体,分布在皮肤真皮乳头内,以手指、足趾的掌侧的皮肤居多,感受触觉,其数量可随年龄增长而减少。触觉小体呈卵圆形,长轴与皮肤表面垂直,外包有结缔组织囊,小体内有许多横列的扁平细胞。有髓神经纤维进入小体时失去髓鞘,轴突分成细支盘绕在扁平细胞间(图7-22)。
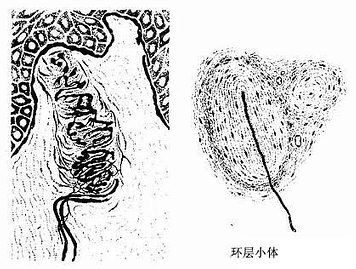
图7-22 有被囊的感觉神经末梢
(2)环层小体(lamellar corpuscle):又称Pacinian小体,体积较大(直径1~4mm),卵圆形或球形,广泛分布在皮下组织、肠系膜、韧带和关节囊等处,感受压觉和振动觉。小体的被囊是由数十层呈同心圆排列的扁平细胞组成,小体中央有一条均质状的圆柱状。有髓神经纤维进入小体失去髓鞘,裸露轴突穿行于小体中央的圆柱体内(图7-22)。
(3)肌梭(muscle spindle):是分布在骨骼肌内的梭形小体,长约1~7mm,外有结缔组织被囊,内含若干条细小的骨骼肌纤维称梭内纤维(intrafusal muscle fiber)。梭内肌纤维的中段肌浆较多,肌原纤维较少,有些肌纤维的细胞核排列成串,有些肌纤维的细胞核聚集在中段而使中段膨大。感觉神经纤维进入肌梭时失去髓鞘,其轴突细支呈环状包绕梭内肌纤维的两端。肌梭是一种本体感受器,主要感受肌纤维的伸缩变化,在调节骨骼肌的活动中起重要作用(图7-23)。

图7-23 肌梭结构模式图
(二)运动神经末梢
运动神经末梢(motor nerve ending)是运动神经元的长轴突分布于肌组织和腺内的终末结构,支配肌纤维的收缩和腺的分泌。神经末梢与邻近组织共同组成效应器(effector)。运动神经末梢又分躯体和内脏运动神经末梢两类。
1.躯体运神经末梢躯体运动神经末梢(somatic motor nerve ending)分布于骨骼肌内。神经元的胞体位于脊髓灰质前角或脑干,轴突很长,离开中枢神经系统后成为躯体传出(运动)神经纤维,其中小部分细有髓神经纤维供应肌梭内的梭内肌纤维,其余大部分粗有髓神经纤维均分布于骨骼肌(梭外肌)。有髓神经纤维抵达骨骼肌时失去髓鞘,其轴突反复分支,每一分支形成葡萄状终末与一条骨骼肌纤维建立突触连接,此连接区域呈椭圆形板状隆起,称运动终板(motor end plate)或神经肌连接(neuromuscular junction)(图7-24)。

图7-24 运动终板(骨骼肌纤维压片,氯化金法)
一条有髓运动神经纤维支配的骨骼肌纤维数目多少不等,少者1~2条,多者可分支支配上千条;而一条骨骼肌纤维通常只有一个轴突分支支配。一个运动神经元的轴突及其分支所支配的全部骨骼肌纤维合称一个运动单位(motor unit)。在电镜下,运动终板处的肌纤维含丰富的肌浆,有较多的细胞核和线粒体,肌纤维表面凹陷成浅糟,突终末嵌入浅糟内(图7-25)。糟底肌膜即突触后膜,它又凹陷成许多深沟和皱褶,使突触后膜的表面积增大,突触后膜上有乙酰胆碱N型受体。轴突终末与肌膜之间在的间隙为突触间隙,与肌膜相对的轴膜是突触前膜,它富含电位门控钙通道。轴突终末内有大量圆形突触小泡,内含神经递质乙酰胆碱。还有许多线粒体和一些微管、微丝等。当神经冲动到达运动终时,轴突终末突触前膜上的电位门控钙通道开放,Ca2+进入轴突终末内,促使突触小泡移附于突触前膜,并藉出胞作用释放其内的乙酰胆碱到突触间隙。大部分乙酰胆碱分子与肌膜(突触前膜)上的乙酰胆碱N型受体结合,引起与受体偶联的化学门控钠通道开放,大量Na+进入肌浆内,使肌膜两侧离子分布发生变化而产生兴奋,从而引起肌纤维的收缩。

图7-25 运动终板超微结构模图
2.内脏运动神经末梢内脏运动神经末梢(visceral motor nerve ending)分布于内脏及心血平滑肌、心肌和腺上皮细胞等处。内脏运动神经属植物神经系统的一部分,它从中枢到效应器的通路一般由两个神经元组成。第一个神经元称节前神经元(preganglionic neuron),胞体位于脊髓灰质侧角或脑干,轴突称节前纤维。第二个神经元称节后神经元(postganglionic neuron),胞体位于植物神经节可神经丛,轴突组成节后纤维。节前纤维离开中枢进入植物神经节或神经丛,与节后神经元的胞体或树突建立突触连接。节后纤维离开植物神经节或神经丛,分布到内脏及血管的平滑肌、心肌和腺细胞,成为内脏运动神经末梢(图7-26)。这类神经纤维较细,无髓鞘,其轴突终末分支常呈串珠样膨体(varicosity)。它是与效应细胞建立突触的部位。膨体内有许多圆形或颗粒型突触小泡,圆形清亮突触小泡含乙酰胆碱,颗粒型突触小泡含去甲肾上腺素或肽类神经递质。
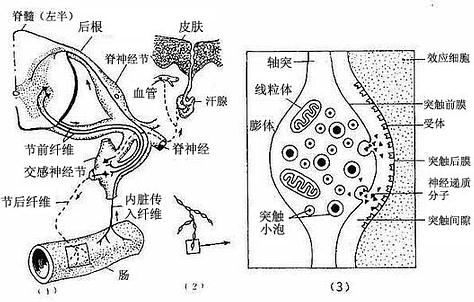
图7-26 内脏运动神经纤维及其末梢
(1)内脏神经分布图解(2)内脏运动神经末梢(3)膨体超微结构图解
根据末梢膨体内突触小泡释放神经递质的不同,可把内脏运动神经纤维分为胆碱能、肾上腺素能和肽能纤维。所有节前纤维都属胆碱能纤维,节后纤维则因交感神经和副交感神经而不同。交感神经的节后纤维是肾上腺素能或肽能纤维,或一种纤维既是肾上腺素能又是肽能的;副交感神经的节后纤维则是胆碱能或肽能纤维,或一种纤维既是胆碱能又是肽能的。与内脏运动神经末梢膨体接触的效应的细胞膜(突触后膜)上有相应的神经递质的不同类型受体,膨体释放的神经递质与不同类型受体结合,可引起效应细胞不同的生理效应。
六、神经纤维的溃变与再生
(一)溃变
神经纤维受损伤如神经被切断后,切断处远侧段的神经纤维全长发生溃变(degeneration),轴突和髓鞘碎裂和溶解。与胞体相连的近侧段则发生逆行性溃变,即轴突和髓鞘的断裂溶解由切断处向胞体方向进行,溃变一般只发展到邻近断端的第一侧支终止。神经元的胞体肿胀,细胞核从中央移到胞体边缘,胞质内尼氏体明显减少,故胞质着色浅淡(图7-27)。
(二)再生
神经元胞体是细胞的营养中心,只有在胞体没有死亡的条件下才有纤维再生(regeneration)的可能。胞体约于损伤后第3周开始恢复,胞质内的尼氏体重新出现,胞体肿胀消失,胞核恢复中央位置。胞体的完全恢复约需3~6个月,恢复中的胞体不断合成新的蛋白质及其他产物输向轴突,使残留的近侧段轴突末端生长出许多新生的轴突支芽(图7-27)。
1.周围神经纤维的再生切断处远侧段的周围神经纤维,虽然其轴突和髓鞘发生溃变,但包裹神经纤维的基膜仍保留呈管状。此时施万细胞大量增生,一面吞噬解体的轴突和髓鞘,一面在基膜管内排列成细胞索,靠近断口处的施万细胞还形成细胞桥把两断端连接起来。从近侧段神经纤维轴突末端长出的轴突支芽,越过此施万细胞桥,进入基膜管内,当其中一支沿着施万细胞索生长并到达原来神经纤维末梢所在处,则再生成功(图7-27)。施万细胞和基膜对轴突的再生起重要的诱导作用。
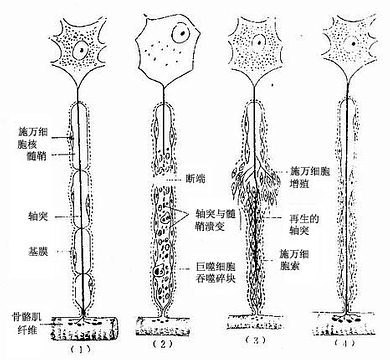
图7-27 周围神经的溃变与再生图解
(1)正常神经纤维,(2)神经纤维断离处及端及近端的一部分髓鞘及轴突溃变(3)施万细胞增生,轴突生长,(4)多余的轴突消失,神经纤维再生完成。
2.中枢神经纤维的再生中枢神经纤维的再生比周围神经困难。神经纤维无施万细胞,亦无基膜包裹。当中枢神经纤维受损伤时,星形胶质细胞增生肥大,在损伤区形成致密的胶质瘢痕,大多数再生轴突支不能越过此胶质瘢痕;即使能越过,也没有如同周围神经纤维那样的基膜管和施万细胞索引导再生轴突到达目的地。所以,中枢神经纤维的损伤常导致脊髓或脑功能的永久性丧失。数十年来不少科学家为研究神经再生进行不懈的努力,近年来已注意到一类能促进神经生长的化学物质称神经营养因子(neurotrophic factor)的作用。同时又根据胚胎神经元容易生长的化学物质称神经营养因子(neurotrophic factor)的作用。同时又根据胚胎神经元容易生长及周围神经能再生的特点,把胚胎脑组织、周围神经或周围神经的组分(如基膜或基膜的化学成分)移植到脑内,以期促进中枢神经再生。
七、神经节、脊髓、大脑皮质和小脑皮质的结构
在中枢神经系统,神经元胞体集中的部分称灰质(gray matter),不含胞体只有神经纤维的部分称白质(white matter)。大脑和小脑的灰质位于脑的表层,故又称皮质(cortex),皮质下是白质。在脑的白质内,神经元胞体集中而成的一些团块称神经核(团)。脊髓的灰质则位于中央,周围是白质。在周围神经系统,神经元胞体集中的部分构成神经节或神经丛。分布在各部位的神经元,它们的形态、大小及突起的长短有很大的差异,其中的长轴突(Golgi型)神经元,其轴突从胞体所在的区域远伸到神经系统的其他部分,或分布到身体的其他组织。
(一)神经节
神经节可服脑脊神经节(cerebrospinal ganglion)和植物神经节(vegetative ganglion)两大类。脑脊神经节位于脊神经后根和某些脑神经干上,植物神经节包括交感和副交感神经节。交感神经节位于脊柱两旁及前方,副交感神经节则位于器官附近或器官内。神经节一般为卵圆形,与周围神经相连,外包结缔组织被膜。节内的神经细胞称节细胞,细胞的胞体被一层扁平的卫星细胞包裹,卫星细胞外面还有一层基膜。除节细胞外,节内还有大量神经纤维以及少量结缔组织和血管。
1.脑脊神经节 属感觉神经节。节细胞是假单极神经元,胞体圆或卵圆形,大小不等,大者直径达100μm以上,小者仅15μm。细胞核圆形位于胞体中央,核仁明显。胞质内的尼氏体细小分散。从胞体发出一个突起,在胞体附近盘曲,然后呈“T”形分支,一支走向中枢(中枢突),另一支(周围突)经脑脊神经分布到外周组织,其末梢形成感受器。卫星细胞包裹着节细胞胞体及其盘曲的突起,在“T”形分支处与施万细胞鞘相连续。节细胞的胞体大多集中在神经节的周缘,并被神经纤维束分隔成群。脑脊神经节内的神经纤维大部分是有髓神经纤维(图7-28)
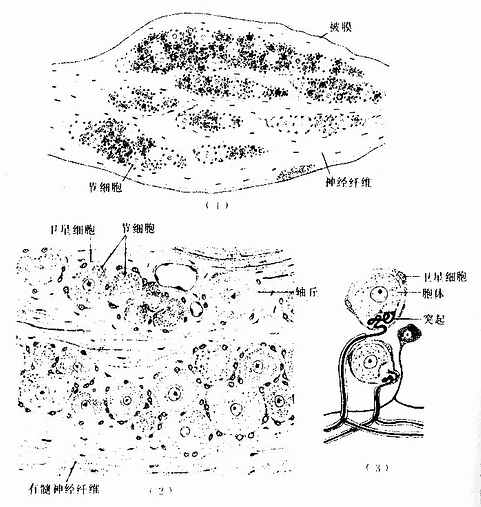
图7-28 脊神经节
(1)低倍 (2)高倍 (3)假单极神经元
2.植物神经节 其中的节细胞是植物神经系统的节后神经元,属多极的运动神经元,胞体一般较感觉神经节的细胞小,散在分布(图7-29)。细胞核常偏位于细胞的一侧,部分细胞有双极,胞质内尼氏体呈颗粒状,均匀分布。卫星细胞数量较少,不完全地包裹节细胞胞体。节内的神经纤维多为髓神经纤维,较分散,其中有节前纤维和节后纤维。节前纤维与节细胞的树突和胞体建立突触,节后纤维离开神经节,其末梢分布到内脏及心血管的平滑肌、心肌和腺上皮细胞,即内脏运动神经末梢。
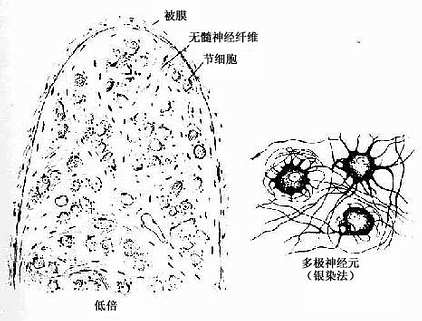
图7-29 交感神经节
交感神经节的节细胞有两种。一种是体积略大的主节细胞(principal ganglion cell),占大多数。大部分主节细胞属肾上腺素能神经元,少数为胆碱能神经元。另一种节细胞数量少,体积也小,常聚集成小群,荧光组织化学染色标本,这些细胞显示强荧光,故称小强荧光细胞(small intensely fluorescenecell,SIF细胞)。SIF细胞释放神经递质多巴胺,它可能是一种中间神经元,其轴突终末与主节细胞建立突触。副交感神经节的节细胞一般属胆碱能神经元。近年来认为,植物神经节除含肾上腺素能和胆碱能神经元外,还存在释放肽类神经递质的肽能神经元。
(二)脊髓灰质
脊髓的灰质居中央,在横切面上呈蝴蝶形(图7-3),分前角、后角和侧角(侧角主要见于胸腰段脊髓)。神经元都是多极型。前角内大多是躯体运动神经元,胞体大小不等。大者称α神经元,其轴突较粗,分布到骨骼肌(梭外肌);小者称γ神经元,其轴突较细,支配梭的梭内肌纤维。还有一种短轴突的小神经元称Ranshaw细胞,其轴突与α神经元的胞体形成突触,可能通过释放甘氨酸,抑制α神经元的活动。侧角内的神经元是交感神经系统的节前神经元,胞体中等大小,其轴突(节前纤维)终止于交感神经节,与节细胞建立突触(图7-26)。前角的躯体运动神经元和侧角的内脏运动神经元都是乙酰胆碱能神经元。后角内的神经元组成较复杂,细胞一般较小,它们主要接受后根纤维(感觉神经元的中枢突)传入的神经冲动,其轴突在白质内形成各种上行纤维束到脑干、小脑和丘脑,所以这类神经元又称为束细胞(tract cell)。此外,脊髓灰质内还遍布许多中间神经元,它们的轴突长短不一,但都离不开脊髓,短轴突与同节段的束细胞和运动神经元联系,长轴突在白质上下穿行至相邻或较远的脊髓节段,终止于同侧或对侧的神经元。
(三)大脑皮质
1.大脑皮质的神经元类型大脑皮质的神经元都是多极神经元,按其细胞的形态分为锥体细胞、颗粒细胞和梭形细胞三大类(图7-30)。
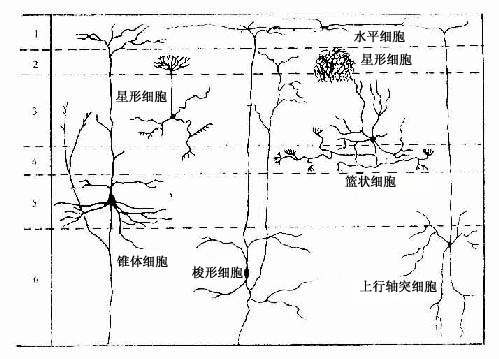
图7-30大脑皮质神经元的形态和分布
(1)锥体细胞(pyramidal cell):数量较多,可分大、中、小三型。胞体形似锥形,尖端发出一条较粗的主树突,伸向皮质表面,沿途发出许多小分支,胞体还向四周发出一些水平走向的树突。轴突自胞体底部发出,长短不一,短者不越出所在皮质范围,长者离开皮质,进入髓质(白质),组成投射纤维(下行至脑干或脊髓)或联合纤维(到同侧或对侧的另一皮质区)。因而,锥体细胞是大脑皮质的主要投射(传出)神经元。
(2)颗粒细胞(granular cell):数目最多。胞体较小,呈颗粒状,包括星形细胞(stellate cell)、水平细胞(horizontal cell)和篮状细胞(basket cell)等几种。以星形细胞最多,它们的轴突多数很短,终止于附近的锥体细胞或梭形细胞。有些星形细胞的轴突较长,上行走向皮质表面,与锥体细胞顶树突或水平细胞相联系。水平细胞的树突和轴突与皮质表面平行分布,与锥体细胞顶树突联系。所以,颗粒细胞是大脑皮质区的局部(中间)神经元,构成皮质内信息传递的复杂微环路。
(3)梭形细胞(fusiform cell):数量较少,大小不一。大梭形细胞也属投射神经元,主要分布在皮质深层,胞体梭形,树突自细胞的上、下两端发出,上端树突多达皮质表面。轴突自下端树突的主干发出,进入髓质,组成投射纤维或联合纤维。
2.大脑皮质的分层 大脑皮质的这些神经元是以分层方式排列的,除大脑的个别区域外,一般可分为6层,从表面至深层的结构如下(图7-31):
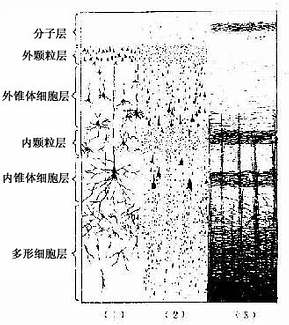
图7-31 大脑皮质6层结构
(1)银染法示神经元形态 (2)尼氏染色示6层结构
(3)髓鞘染色示神经纤维的分布
(1)分子层(molecular layer):神经元小而少,主要是水平细胞和星形细胞,还有许多与皮质表面平行的神经纤维。
(2)外颗粒层(external granular layer):主要由许多星形细胞和少量小型锥体细胞构成。
(3)外锥体细胞层(external pyramidal layer):此层较厚,由许多中、小型锥体细胞和星形细胞组成。
(4)内颗粒层(internal granular layer):细胞密集,多数是星形细胞。
(5)内锥体细胞层(internal pyramidal layer):主要由中型和大型锥体细胞组成。在中央前回运动区,此层有巨大锥体细胞,胞体高120μm,宽80μm,称Betz 细胞,其顶树突伸到分子层,轴突下行到脑干和脊髓。
(6)多形细胞层(polymorphic layer):以梭形细胞为主,还有锥体细胞和颗粒细胞。
大脑皮质的1-4层主要接受传入冲动。从丘脑来的特异传入纤维(各种感觉传入的上行纤维)主要进入第4层与星形细胞形成突触,星形细胞的轴突又与其他细胞建立广泛的联系,从而对传入皮质的各种信息进行分析,作了反应。起自大脑半球同侧或对侧的联合传入纤维则进入第2、3层,与锥体细胞形成突触。大脑皮质的传出纤维分投射纤维和联合纤维两种。投射纤维主要起自第5层的锥体细胞和第6层的大梭形细胞,下行至脑干及脊髓。联合纤维起自第3、5、6层的锥体细胞和梭形细胞,分布于皮质的同侧及对侧脑区。皮质的第2、3、4层细胞主要与各层细胞相互联系,构成复杂的神经微环路对信息进行分析、整合和贮存(图7-32)。大脑的高级神经活动可能与其复杂的微环路有密切关系。
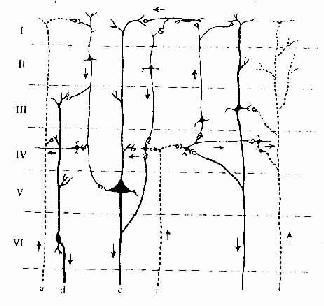
图7-32大脑皮质内微环路简图
a 特异传入纤维,b联合传入纤维,c 投射传出纤维,d联合传出纤维
大脑皮质的6层结构因不同脑区而有差异。例如中央前回(运动同)的第4层不明显,第5层较发达,有巨大锥体细胞(细胞);视皮质则第4层特别发达,第5层的细胞较小。有些学者对大脑皮质进行了组织学普查,根据细胞的排列和类型以及有髓神经纤维的配布型式等的差异,作出了人脑皮质的分区图。不同的学者有不同的分区法,较常用的是Brodmann(1909)的分区法,把大脑皮质分为52区,并以数字表示之。
虽然大脑皮质的神经元是以分层方式排列的,但对大脑皮质结构与功能的研究发现,皮质细胞是呈纵向柱状排列的,称此为垂直柱(vertical column)。垂直柱可能是构成大脑皮质的基本功能单位,如感觉皮质区的一个垂直柱内的神经元。具有相同或近于相同的周围感受野,即这些神经元都对同一类型的周围刺激起反应。同垂直柱贯穿皮质全厚,大小不等,直戏约350~450μm,它包括传入纤维,传出神经元和中间神经元,传入纤维直接或间接通过柱内各层细胞构成复杂的反复回路,然后作用于同的传出神经元。皮质垂直柱内除垂直方向的反复回路外,还可通过星形细胞和锥体细胞的基底树突使兴奋横向扩布,影响更多垂直柱的神经元活动。
(四)小脑皮质
小脑表面有许多平行的横沟,把小脑分隔成许多小叶片。每一叶片表面是一层灰质,即小脑皮质,皮质下为白质(髓质)。小脑皮质从外到内明显地分3层(图7-33)。皮质内的神经元有星形细胞、篮状细胞、蒲肯野细胞(Purkinje cell或称梨状细胞)、颗粒细胞和高尔基细胞(Golgi cell)5种(图7-34)。
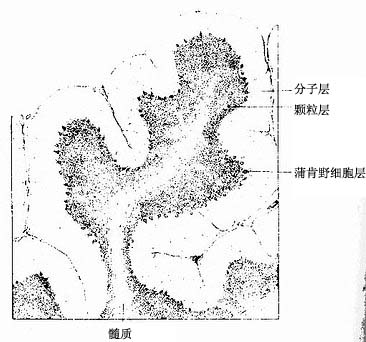
图7-33 小脑切面
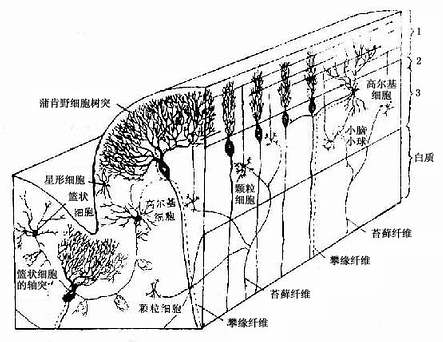
图7-34 小脑皮质结构图解
1.分子层 2.蒲肯野细胞层 3.颗粒层
1.分子层此层较厚,神经元较少,主要有两种。一种是小型多突的星形细胞,轴突较短,分布于浅层。另一种是篮状细胞,胞体较大,分布于深层,其轴突较长,与小脑叶片长轴成直角方向并平行于小脑表面走行,沿途发出许多侧支,其末端呈篮状分支包绕蒲肯野细胞的胞体并与之形成突触。
2.蒲肯野细胞层由一层蒲肯野细胞胞体组成。此种细胞是小脑皮质中最大的神经元,胞体呈梨形,从顶端发出2~3条粗的主树突伸向分子层,树突的分支繁多,形如侧柏叶状或扇形,铺展在与小脑叶片长轴垂直的平面上。轴突自胞体底部发出,离开皮质进入髓质,终止于小脑内部的核群。
3.颗粒层 此层由密集的颗粒细胞和一些高尔基细胞组成。颗细胞很小,胞体直径与淋巴细胞近似,有4~5个短树突,树突末端分支如爪状。轴突上行进入分子层呈“T”形分支,与小脑叶片长轴平行,称平行纤维(parallel fiber)。平行纤维穿过一排排蒲肯野细胞的扇形树突,与其树突棘形成突触(图7-35)。一条平行纤维可与400多个蒲肯野细胞建立突触,每个蒲肯野细胞与一条平行纤维之间只有一个突触连接;但一个蒲肯野细胞的扇形树突有20万~30万条平行纤维通过,故一个蒲肯野细胞的树突上共有20万~30万个突触。高尔基细胞的胞体较大,树突分支较多,大部分伸入分子层与平行纤维接触,轴突在颗粒层内呈丛密分支,与颗粒细胞的树突形成突触。
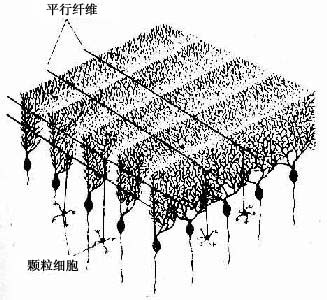
图7-35 蒲肯野细胞的排列模式与平行纤维的关系
小脑的5种神经元中,蒲肯野细胞是唯一的传出神经元。颗粒细胞是谷氨酸能的兴奋性神经元,其他中间神经元都是γ-氨基丁酸(GABA)能的抑制性神经元。小脑的传入纤维有三种:攀缘纤维(climbing fiber)、苔藓纤维(mossy fiber)和去甲肾上腺素能纤维。前两者是兴奋性纤维,后者是抑制性纤维。攀缘纤维主要起源于延髓的下橄榄核,纤维较细,它进入小脑皮质攀附在蒲肯野细胞的树突上与之形成突触。一条攀缘纤维与一个蒲肯野细胞树突所形成的突触可达300多个。故一条攀缘纤维的神经冲动可引起一个蒲肯野细胞强烈兴奋。苔藓纤维起源于脊髓和脑干的核群,纤维较粗,进入小脑皮质后纤维末端分支呈苔状。每一膨大的末端可与许多(约20个)颗粒细胞的树突、高尔基细胞的轴突或近端树突形成复杂的突触群,形似小球,故称小脑小球(cerebellar glomerulus)(图7-36)。一条苔藓纤维的分支可分布于2个或2个以上的叶片内,约可兴奋800多个颗粒细胞,每个颗粒细胞的平行纤维又与400多个蒲肯野细胞接触。这样,一条苔藓纤维可引起几十万个蒲肯野细胞兴奋,所以,攀缘纤维和苔藓纤维把来自小脑外的神经冲动传到小脑皮质,最后都作用于蒲肯野细胞。攀缘纤维直接强烈地兴奋单个蒲肯野细胞,而苔藓纤维则通过颗粒细胞的平行纤维间接兴奋几十万个蒲肯野细胞。另一方面,攀缘纤维的侧支及颗粒细胞的平行纤维还可以与其他抑制性中间神经元(星形细胞、篮细胞和高尔基细胞)形成突触,这些抑制性中间神经元又与肯野细胞形成突触。因此,攀缘纤维的冲动可通过其侧支作用于抑制性中间神经元,从而抑制蒲肯野细胞。同样,苔藓纤维通过颗粒细胞平行纤维兴奋许多蒲肯野细胞的同时,亦可通过与抑制性中间神经元连接,抑制蒲肯野细胞的兴奋。此外,去甲肾上腺素能纤维(来自脑干的蓝斑核)对蒲肯野细胞亦有抑制作用。
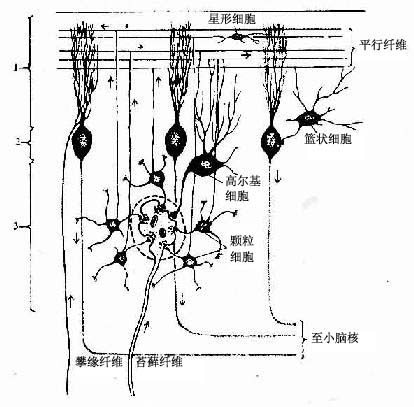
图7-36 小脑皮质神经元与传入纤维的关系
1分子层 2蒲肯野细胞层 3颗粒层
虚线范围代表一个小脑小球
八、脑脊膜和血脑屏障
脑脊膜是包在脑和脊髓外面的结缔组织膜,有三层,由外向内是硬膜(dura mater)、蛛网膜(arachnoid )和软膜(pia mater)。硬膜是较厚而坚韧的致密结缔组织,其内表面有一层间皮细胞覆盖。硬膜与蛛网膜之间有一狭窄的间隙,称硬膜下隙(subdural space),内含少量液体。蛛网膜是由薄层纤细的结缔组织构成,它与软膜之间有较宽大的腔隙称蛛网膜下隙(subarachnoid space)。蛛网膜的结缔组织纤维形成许多小梁与软膜相连,小梁在蛛网膜下隙内分支形成蛛网膜结构。蛛网膜下隙内含脑脊液。软膜是紧贴在脊髓表面的薄层结缔组织,富含血管。在软膜外表面和蛛网膜的外、内表面以及小梁的表面均被覆有单层扁平上皮,软膜的血管供应脑及脊髓。血管进入脑内时,软膜和蛛网膜也随之进入脑内,但软膜并不紧包血管,血管与软膜之间仍有空隙,称血管周隙(perivascular space),与蛛网膜下隙相通,内含脑脊液(图7-37)。当小血管进一步分支形成毛细血管时,软膜组织和血管周隙都消失,毛细血管则由星形胶质细胞突起所包裹(图7-37、7-14)。
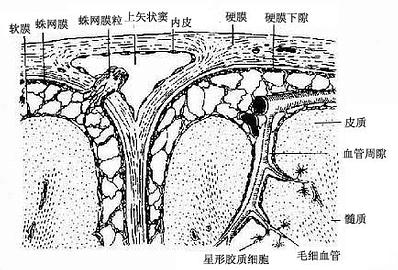
图7-37 大脑冠状切面,示脑膜和血管
脑的毛细血管与身体其他器官的毛细血管不同,它能阻止多种物质进入脑。例如将一种活性染料台盼蓝(trypan blue)注入动物的血内,身体各器官都被染上蓝色,唯独脑组织不着色。因此认为血液与脑组织之间在存在一种血脑屏障(blood-brain barrieer,BBB),限制某些物质进入脑组织。此血脑屏障由脑毛细血管内皮细胞、基膜和神经胶质膜构成。脑的毛细血管属连续型(见循环系统),毛细血管内皮细胞之间以紧密连接封闭,内皮外有基板、周细胞及星形胶质细胞突起的脚板围绕(图7-38)。实验证明,内皮细胞是构成血脑屏障的主要结构,它可阻止多种物质进入脑,但营养物质和代谢产物可顺利通过,以维持神经系统内环境的相对稳定。脑毛细血管内皮细胞的这种生理特性,与细胞膜上存在许多不同类型的转运器(transporter)有关,转运器是细胞膜上的蛋白质,能识别特定分子并转运它们越过血脑屏障。星形胶质细胞有诱导内皮细胞形成屏障特性的作用。
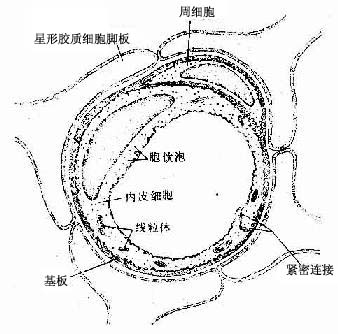
图7-38 脑毛细血管超微结构模式图
九、脉络丛和脑脊液
脉络丛(choroid plexus)见于第Ⅲ、Ⅳ脑室顶和部分侧脑室壁,它是由富含血管的软膜与室管膜直接相贴并突入脑室而成的皱襞状结构,室管膜则成为有分泌功能的脉络丛上皮(图7-39)。脉络丛上皮由一层立方形或矮柱形细胞组成,细胞表面有许多微绒毛,细胞核大而圆,胞质内线粒体很多。细胞侧面之间靠近游离面处有连接复合体。上皮下是基膜,基膜深部是结缔组织。结缔组织内含丰富血管和巨噬细胞。毛细血管属有孔型,内皮细胞上的小孔有薄隔膜封闭。

图7-39 脉络丛
脉络丛的主要功能是产生脑脊液(cerebrospinal fluid),脑脊液是由脉络丛上皮细胞分泌的,为无色透明的液体,含蛋白质很少,但有较高浓度的Na+、K+和Cl-,并有少许脱落细胞和淋巴细胞。成年男性约有100ml脑脊液,在脑室、脊髓中央管、蛛网膜下隙和血管周隙。脑脊液通过蛛网膜粒(蛛网膜突入颅静脉窦内的绒毛状突起)吸收入血(图7-37)。脉络丛上皮不断分泌脑脊液,又不断回流入血液,形成脑脊液循环。脉络丛上皮和脉络丛毛细血管内皮共同构成血-脑脊液屏障(blood-cerebrospinal fluidbarrier,BCB),使脑脊液保持稳定的成分而不同于血液。脑脊液有营养和保护脑与脊髓的作用。

 中医世家
中医世家 浦 标 网
浦 标 网 河南大学精品课程
河南大学精品课程 图书资料室
图书资料室