消化系统由消化管与消化腺组成,主要功能是对食物进行物理性和化学性消化,将大分子物质分解为小分子的氨基酸、单糖、甘油酯等。吸收后供机体生长和代谢的需要。消化管是从口腔至肛门的连续性管道,依次分为口腔、咽、食管、胃、小肠和大肠。这些器官的管壁结构具有某些共同的分层规律,又各具有与其功能相适应的特点。
一、消化管的一般结构
消化管(除口腔与咽外)自内向外均分为粘膜、粘膜下层、肌层与外膜四层(图12-1)。
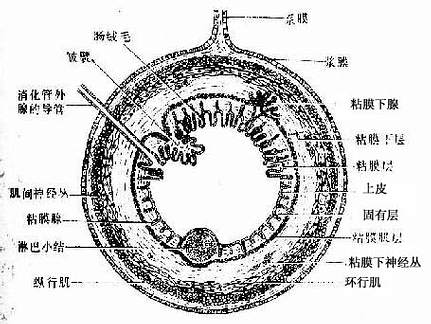
图12-1 消化管一般结构模式图
(一)粘膜
粘膜(tunica mucosa)由上皮、固有层和粘膜肌组成,是消化管各段结构差异最大、功能最重要的部分。
1.上皮 上皮的类型依部位而异。消化管的两端(口腔、咽、食管及肛门)为复层扁平上皮,以保护功能为主;其余部分均为单层柱状上皮,以消化吸收功能为主。上皮与管壁内的腺体相连。
2.固有层 固有层(lamina propria )为疏松结缔组织,含细胞和纤维较多,并有丰富的血管和淋巴管。胃肠固有层内还富含腺体或淋巴组织。
3.粘膜肌层 粘膜肌层(muscularis mucosa)为薄层平滑肌,其收缩可使粘膜活动,促进固有层内的腺体分泌物排出和血液运行,利于物质吸收。
(二)粘膜下层
粘膜下层(tela submucosa)由疏松结缔组织组成,内含较大的血管与淋巴管。在食管及十二指肠的粘膜下层内分别有食管腺与十二指肠膜。粘膜下层中还有粘膜下神经丛,由多极神经元与无髓神经纤维构成,可调节粘膜肌的收缩和腺体的分泌。在食管、胃和小肠等部位的粘膜与粘膜下层共同向管腔内突起,形成皱襞(plica)。
(三)肌层
除食管上段与肛门处的肌层(tunica muscularis)为骨骼肌外,其余大部均为平滑肌。肌层一般分为内环行、外纵行两层,其间有肌间神经丛,结构与粘膜下神经丛相似,可调节肌层的运动。
(四)外膜
外膜(tunica adventitia)由薄层结缔组织构成者称纤维膜(fibrosa),主要分布于食管和大肠末段,与周围组织无明显界限。由薄层结缔组织与间皮共同构成者称浆膜(serosa),见于腹膜内位的胃、大部分小肠与大肠,其表面光滑,利于胃肠活动。
二、口腔
(一)口腔粘膜的一般结构
口腔粘膜只有上皮和固有层,无粘膜肌。上皮为复层扁平,仅在硬腭部出现角化。固有层结缔组织突向上皮形成乳头,其内富有毛细血管,故新鲜粘膜呈红色。乳头及上皮内有许多感觉神经末梢。固有层中尚有粘液性和浆液性的小唾液腺。固有层下连骨骼肌(于唇、颊等处)或骨(于硬腭)。
(二)舌
舌由表面的粘膜和深部的舌肌组成。舌肌由纵行、横行及垂直走行的骨骼肌纤维束交织构成。粘膜由复层扁平上皮与固有层组成。粘膜由复层扁平上皮与固有层组成。舌根部粘膜内有许多淋巴小结,构成舌扁桃体。舌背部粘膜形成许多乳头状隆起,称舌乳头(lingual papillae),可分为四种。
1.丝状乳头 丝状乳头(filiform papillae)数目最多,遍布于舌背各处。乳头呈圆锥形,尖端略向咽部倾斜,浅层上皮细胞角化脱落,外观白色,称舌苔(图12-2)。
2.菌状乳头 菌状乳头(fungif[alt] [/alt][/img]2在碳酸酐酶作用下与H2O结合形成H2CO3;H2CO3解离为H+和HCO3-,H+被主动运输至分泌小管,而HCO3-与血液中的CL-次换;CL-也被运输入分泌小管,与H+结合成盐酸(图12-11)。盐酸能激活胃蛋白酶原,使之成为胃蛋白酶,对蛋白质进行初步分解;盐酸还有杀菌作用。人的壁细胞还分泌内因子(intrinsic factor),这种糖蛋白在胃腔内与食物中的维生素B12结合成复合物,使 B12在肠管内不被酶分解,并能促进回肠吸收B12入血,供红细胞生成所需。如内因子缺乏,维生素B12吸收障碍,可导致恶性贫血。
颈粘液细胞(nedk mucous cell):数量很少,位于腺颈部,多呈楔形夹于其它细胞间。核多呈扁平形,居细胞基底,核上方有很多粘原颗粒,HE染色浅淡,故常不易与主细胞相区分,其分泌物为含酸性粘多糖的可溶性粘液。
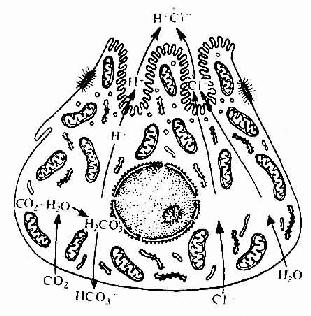
图12-11 壁细胞合成盐酸示意图
内分泌和细胞:见后述。
主细胞和壁细胞的寿命为200余天,衰老的细胞在胃底腺底部脱落,新增殖的细胞从颈部向底部缓慢迁移。由于在颈部尚未发现典型的未分化细胞,故目前一般认为颈粘液细胞可分化为其它胃底腺细胞;主细胞自身也具有一定的分裂能力。
(2)贲门腺(cardiac gland):分布于近贲门处宽5~30mm的狭窄区域,为分支管状的粘液腺,可有少量壁细胞。
(3)幽门腺(phloric gland):分布于幽门部宽4~5cm的区域,此区胃小凹甚深。幽门腺为分支较多而弯曲的管状粘液腺,内有较多内分泌细胞。
3.粘膜肌层由内环行与外纵行两层平滑肌组成。内环肌的部分细胞伸入固有层腺体之间,其收缩有助于腺分泌物的排出。
胃粘膜的自我保护机制:胃液含高浓度盐酸,PH值为2,腐蚀力极强,胃蛋白酶能分解蛋白质,而胃粘膜却不受破坏,这主要是由于胃粘膜表面存在粘液-碳酸氢盐屏障(mucous-HCO3- barrier)。胃上皮表面覆盖的粘液层厚0.25~0.5mm ,主要由不可溶性粘液凝胶(mucingel)构成,并含大量HCO3-,后者部分由表面粘液细胞产生,部分来自壁细胞(图12-12)。凝胶层将上皮与胃蛋白酶相隔离,并减缓H+向粘膜方向的弥散;HCO-3可中和H+,形成H2CO3,后者被胃上皮细胞的碳酸酐酶迅速分解为H2O和CO2。此外,胃上皮细胞的快速更新也使胃能及时修复损伤。
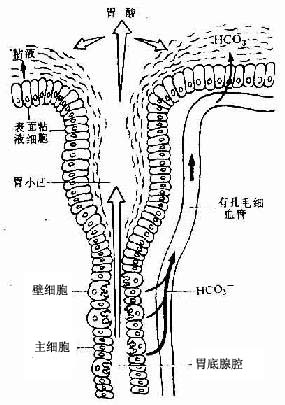
图12-12 胃粘液-碳酸氢盐屏障示意图
(二)粘膜下层
为疏松结缔组织,内含较粗的血管、淋巴管和神经,尚可见成群的脂肪细胞。
(三)肌层
较厚,一般由内斜行、中环行及外纵行三层平滑肌构成。环形肌在贲门和幽门部增厚,分别形成贲门括约肌和幽门括约肌。
(四)外膜
为浆膜。
六、小肠
小肠是消化和吸收的主要部位,分为十二指肠、空肠和回肠,各具某些结构特点。
(一)粘膜
小肠腔面的环行皱襞从距幽门约5cm处开始出现,在十二指肠末段和空肠头段极发达,向下逐渐减少和变矮,至肠中段以下基本消失。粘膜表面还有许多细小的肠绒毛(intestinal villus),是由上皮和固有层向肠腔突起而成,长0.5~1.5mm,形状不一,以十二指肠和空肠头段最发达(图12-13,12-14)。绒毛于十二指肠呈叶状,于空肠如指状,于回肠则细而短(图12-15)。环行皱襞和绒毛使小肠表面积扩大20~30倍,总面积达20m2左右。绒毛根部的上皮下隐至固有层形成管状的小肠腺(small intestinal gland),又称肠隐窝(intestinal crypt),故小肠腺与绒毛的上皮是连续的,小肠腺直接开口于肠腔。
图12-13 大鼠十二指肠绒毛扫描电镜像×230
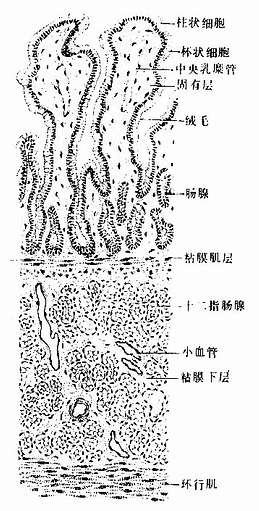
图12-14 十二指肠(横切)
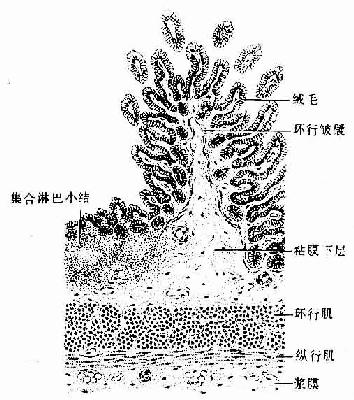
图12-15 回肠(纵切)
1.上皮为单层柱状。绒毛部上皮由吸收细胞、杯状细胞和少量内分泌细胞组成;小肠腺上皮除上述细胞外,还有潘氏细胞和未分化细胞。
(1)吸收细胞(absorptive cell):最多,呈高柱状,核椭圆形,位于细胞基部。绒毛表面的吸收细胞游离面在光镜下可见明显的纹状缘,电镜观察表明它是由密集而规则排列的微绒毛构成(图12-16)。每个吸收细胞约有微绒毛1000根,每根长1~1.4μm ,粗约80nm ,使细胞游离面面积扩大约20倍。小肠腺的吸收细胞的微绒毛较少而短,故纹状缘薄。微绒毛表面尚有一层厚0.1~0.5μm的细胞衣,它是吸收细胞产生的糖蛋白,内有参与消化碳水化合物和蛋白质的双糖酶和肽酶,并吸附有胰蛋白酶、胰淀粉酶等,故细胞衣是消化吸收的重要部位。微绒毛内有纵行微丝束,它们下延汇入细胞顶部的终末网(图12-17)。吸收细胞胞质内有丰富的线粒体和滑面内质网。滑面内质网膜含有的酶可将细胞吸收的甘油-酯与脂肪酸合成甘油三酯,后者与胆固醇、磷脂及β-脂蛋白结合后,于高尔基复合体形成乳糜微粒,然后在细胞侧面释出,这是脂肪吸收与转运的方式。相邻细胞顶部之间有紧密连接、中间连接等构成的连接复合体,可阻止肠腔内物质由细胞间隙进入组织,保证选择性吸收的进行(图12-17)。

图12-16 小鼠小肠吸收细胞电镜像 ×9900
Mv微绒毛,N细胞核
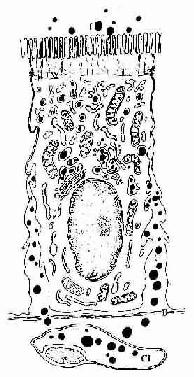
图12-17 小肠吸收细胞
f脂肪 TW终末网 S滑面内质网 G高尔基复合体
C乳糜微粒B基膜 CL中央乳糜管
(2)杯状细胞(goblet cell): 散在于吸收细胞间,分泌粘液,有润滑和保护作用,从十二指肠至回肠末端,杯状细胞逐渐增多。
(3)潘氏细胞(Paneth cell):是小肠腺的特征性细胞,位于腺底部,常三五成群。细胞呈锥体形,胞质顶部充满粗大嗜酸性颗粒,内含溶菌酶等,具有一定的灭菌作用(图12-18)。
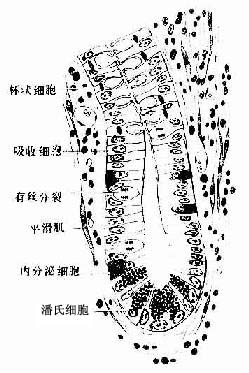
图12-18 小肠腺模式图
(4)内分泌细胞(见后述)。
(5)未分化细胞(undifferentiated cell):位于小肠腺下半部,散在于其它细胞之间。胞体较小,呈柱状,胞质嗜碱性。细胞不断增殖、分化、向上迁移,以补充绒毛顶端脱落的吸收细胞和杯状细胞。绒毛上皮细胞的更新周期为2~4天。一般认为,内分泌细胞和潘氏细胞亦来源于未分化细胞。
2.固有层在细密的结缔组织中除有大量小肠腺外,还有丰富的游走细胞,如淋巴细胞、浆细胞、巨噬细胞,嗜酸性粒细胞等。绒毛中轴的固有层结缔组织内有1~2条纵行毛细淋巴管,称中央乳糜管(central lacteal),它的起始部为盲端,向下穿过粘膜肌进入粘膜下层形成淋巴管丛。中央乳糜管管腔较大,内皮细胞间隙宽,无基膜,故通透性大。吸收细胞释出的乳糜微粒入中央乳糜管输出。此管周围有丰富的有孔毛细血管网,肠上皮吸收的氨基酸、单糖等水溶性物质主要经此入血。绒毛内还有少量来自粘膜肌的平滑肌纤维,可使绒毛收缩,利于物质吸收和淋巴与血液的运行。
固有层中除有大量分散的淋巴细胞外,尚有淋巴小结。在十二指肠和空肠多为孤立淋巴小结,在回肠多为若干淋巴小结聚集形成的集合淋巴小结,它们可穿过粘膜肌抵达粘膜下层(图12-15;见后述)。
3.粘膜肌层 由内环行与外纵行两层平滑肌组成。
(二)粘膜下层
为疏松结缔组织,含较多血管和淋巴管。十二指肠的粘膜下层内有十二指肠腺(duodenal gland),为复管泡状的粘液腺,其导管穿过粘膜肌开口于小肠腺底部(图12-14)。此腺分泌碱性粘液(pH8.2~9.3),可保护十二指肠粘膜免受酸性胃液的侵蚀。最近研究表明,人十二指肠腺尚分泌尿抑胃素(urogasterone),释入肠腔,具有抑制胃酸分泌和刺激小肠上皮细胞增殖的作用。
(三)肌层
由内环行与外纵行两层平滑肌组成。
(四)外膜
除十二指肠后壁为纤维膜外,小肠其余部分均为浆膜。
七、大肠
大肠分为盲肠(附阑尾)、结肠和直肠,主要功能是吸收水分和电解质,将食特残渣形成粪便。大肠粘膜不形成环行皱襞与绒毛,故表面平滑;但在结肠袋之间的横沟处,结肠壁内面有半月形皱襞。
(一)盲肠与结肠
1.粘膜上皮是单层柱状,由柱状细胞和杯状细胞组成,后者数量明显多于小肠。固有层内有大量由上皮下陷而成的大肠腺(亦称肠隐窝),呈长单管状,除含柱状细胞、杯状细胞外,尚有少量未分化细胞和内分泌细胞,无潘氏细胞。固有层内有散在的孤立淋巴小结。粘膜肌层同小肠(图12-19)。
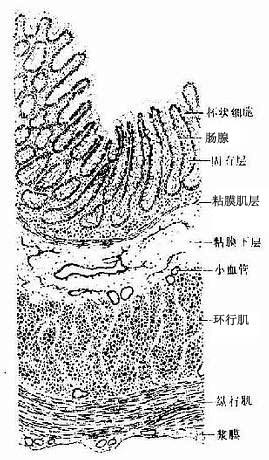
图12-19 结肠(纵切)
2.粘膜下层 在疏松结缔组织内有较大的血管和淋巴管,有成群的脂肪细胞。
3.肌层 为内环行与外纵行两层平滑肌组成。内环行肌较规则,外纵行肌局部增厚形成三条结肠带,带间的纵行肌很薄。
4.外膜 在盲肠、横结肠、乙状结肠为浆膜;在升结肠与降结肠的前壁为浆膜,后壁为纤维膜。外膜结缔组织中常有脂肪细胞集聚构成的肠脂垂。
(二)阑尾
阑尾的管腔小而不规则,大肠腺短而少。固有层内有极丰富的淋巴组织,形成许多淋巴小结,并突入粘膜下层,致使粘膜肌很不完整。肌层很薄,外覆浆膜(图12-20)
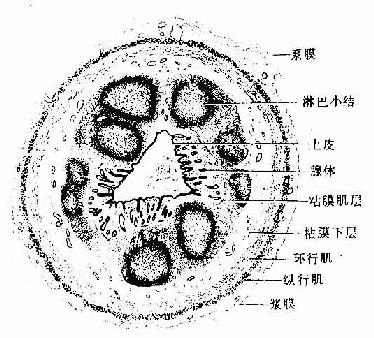
图12-20 阑尾(横切)
(三)直肠
在齿状线以上的直肠粘膜结构与结肠相似。在齿状线处,单层柱状上皮骤变为未角化的复层扁平上皮,大肠腺与粘膜肌消失(图12-21)。痔环以下为角化的复层扁平上皮,近肛门处有环肛腺(顶泌汗腺)。粘膜下层的结缔组织中有丰富的静脉丛,如静脉瘀血扩张则形成痔。肌层为内环行、外纵形两层平滑肌、内环行肌在直肠下段的肛管处增厚形成肛门内括约肌。近肛门处,外纵行肌周围有骨骼肌形成的肛门外括约肌。外膜于直肠上1/3段的大部,中1/3段的前壁为浆膜,其余部分为纤维膜。
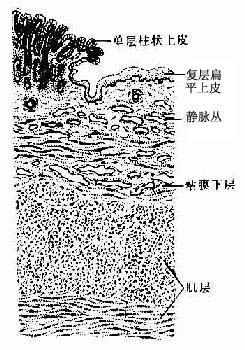
图12-21 直肠齿状线部
八、消化管的淋巴组织及其免疫功能
消化管与机体外环境相通连,各种细菌、病毒、寄生虫卵等有害抗原物质不可避免地随饮食进入。它们大多被胃酸和消化酶所破坏,其余或以原形排出体外,或受到消化管淋巴组织的免疫抵御。消化管淋巴组织又称肠相关淋巴组织(gut- associated lymphoid tissue),包括粘膜淋巴小结(尤以咽、回肠与阑尾处发达),固有层中弥散分布的淋巴细胞、浆细胞、巨噬细胞,上皮内的淋巴细胞等成分。消化管淋巴组织能接受消化管内的抗原刺激,并主要通过产生和向消化管腔分泌免疫球蛋白作为应答。
在肠集合淋巴小结处,局部粘膜向肠腔呈圆顶状隆起,无绒毛和小肠腺。此部位上皮内有散在的小结相关上皮细胞(follicular associated cell),因其游离面有一些微皱褶与短小的绒毛,故又称微皱褶细胞(microfold cell,M细胞)。M细胞基底面质膜内陷形成一较大的穹窿状凹腔,凹腔内含有一至多个淋巴细胞。M细胞下方的基膜多不完整,淋巴细胞易通过。M细胞在光镜下难分辨,只能根据其基底部是否包含淋巴细胞来推断。电镜下可见M细胞胞质很少,但有较多线粒体和丰富的囊泡。这些囊泡被认为是M细胞转运抗原物质的形式。M细胞可摄取肠腔内的抗原物质,并将其传递给下方的淋巴细胞。后者进入粘膜淋巴小结与肠系膜淋巴结内分化增殖,然后经淋细胞再循环途经大部分返回肠粘膜,并转变为浆细胞。浆细胞除产生少量免疫球蛋白G(IgG)进入循环系统外,主要产生免疫球蛋白A(IgA)。IgA能与吸收细胞基底面和侧面膜中的一种称为分泌片(sectetory piece)的镶嵌糖蛋白相结合,形成分泌性IgA(secretory IgA,sIgA)。sIgA被吸收细胞内吞入胞质,继而释入肠腔(图12-22)。sIgA可特异性地与抗原结合,从而抑制细菌增殖,中和病毒,降低抗原物质与上皮细胞的粘着与进入,保护肠粘膜。部分增殖的淋巴细胞还可经血流至其它器官如呼吸道粘膜、女性生殖道粘膜和乳腺等,发挥相似的免疫作用,使消化管免疫成为全身免疫的一部分。
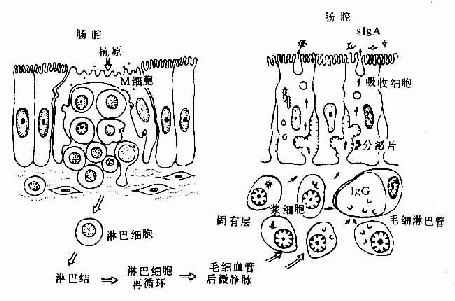
图12-22 消化管粘膜的免疫功能示意图
九、胃肠的内分泌细胞
在胃、小肠与大肠的上皮与腺体中散在着种类繁多的内分泌细胞,其中尤以胃幽门部和十二指肠上段为多。由于胃肠道粘膜的面积巨大,这些细胞的总量超过其它内分泌腺腺细胞的总和。因此,在某种意义上,胃肠是体内最大、最复杂的内分泌器官。它们分泌的多种激素统称胃肠激素(gut hormone),一方面协调胃肠道自身的运动和分泌功能,也参与调节其它器官的活动。
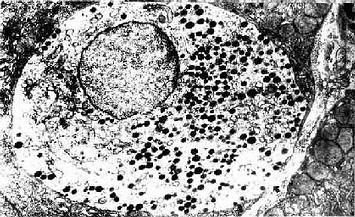
图12-23 小鼠肠腺内的APUD细胞 ×13200
↑基膜 (上海医科大学电镜室供图)
胃肠内分泌细胞大多单个地夹于其它上皮细胞之间,呈不甚规则的圆锥形。基底部附于基膜,并有基底侧突与邻近细胞相接触。胞质中含一些粗面内质网与高尔基复合体。细胞最显著的形态特点是底部胞质中含大量分泌颗粒,故又称基底颗粒细胞(basal granular cell)(图12-23)。分泌颗粒的大小、形状与电子密度依细胞类型而异。绝大部分细胞具有面向管腔的游离面,称开放型,游离面上有微绒毛伸出。此型细胞对管腔食物的刺激和pH变化等化学信息有较强的感受性,从而引起其内分泌活动的变化。少数细胞的顶部被相邻细胞覆盖而未露出腔面,称封闭型,主要受胃肠运动的机械刺激或其它激素的调节而改变其内分泌状态(图12-24)。分泌颗粒中含肽和(或)胺类激素,大多在细胞基底面释入固有层中的毛细血管,经血循环运送并作用于靶细胞;少数激素被释放后可直接作用于邻近细胞,以旁分泌方式调节靶细胞的生理功能。在HE染色切片上,胃肠内分泌细胞不易辩认;用铬或银盐浸染,少数种类的细胞可因其分泌颗粒具嗜铬性、嗜银性或亲银性而被显示。目前主要用免疫组织化学方法来显示这些细胞。
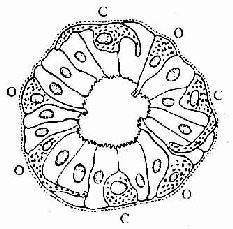
图12-24 消化管内分泌细胞模式图
示开放型(O)与封闭型(C)内分泌细胞
表12-1 胃肠内分泌细胞
| 细胞名称 |
分 布 部 位 |
分 泌 物 |
| 胃 |
肠 |
| D |
胃底、幽门 |
空肠、回肠、结肠 |
生长抑素(somatostatin) |
| D1 |
胃底、幽门 |
空肠、回肠、结肠 |
血管活性肠肽(vasoactive intestinal peptide VIP) |
| EC |
胃底、幽门 |
空肠、回肠、结肠 |
5-羟色胺(5-hydroxytryptamine,5-HT)、P物质(substance P) |
| ECL |
胃底 |
|
组胺(histamine) |
| G |
幽门 |
十二指肠 |
胃泌素(gastrin) |
| I |
|
十二指肠、空肠 |
胆囊收缩素-促胰酶素(cholecystokinin-pancreozymin,CCK-PZ) |
| K |
|
空肠、回肠 |
抑胃多肽(gastric inhibitory polypeptide,GIP) |
| L |
|
空肠、回肠、结肠 |
肠高血糖素(enteroglucagon) |
| M0 |
|
空肠、回肠 |
胃肠动素(motilin) |
| N |
|
回肠 |
神经降压素(neurotensin) |
| P |
胃底、幽门 |
空肠 |
蛙皮素(bombesin) |
| PP |
胃底、幽门 |
结肠 |
胰多肽(pancreatic polypeptide) |
| S |
|
十二指肠、空肠 |
促胰液素(secretin) |
目前已知 余种胃肠内分泌细胞(表12-1),其中研究得比较清楚的有:①EC细胞,数量最多,分布广泛,在胃与空肠尤其丰富。分泌的5-羟色胺可刺激平滑肌收缩,与肠的运动有关,,尚可抑制胃酸分泌,扩张血管。②ECL细胞,仅分布于胃底腺,释放的组胺主要作用于邻近的壁细胞,刺激盐酸分泌。③G细胞,主要分布于胃幽门部,分泌的胃泌素对壁细胞的泌酸功能有强烈的刺激作用。④I细胞,多见于十二指肠和空肠,产生的激素兼有促进胰外分泌部的胰酶分泌和胆囊收缩的作用,故称为胆囊收缩素-促胰酶素。⑤S细胞,亦主要分布于十二指肠和空肠,产生的促胰液素可刺激胰导管上皮细胞分泌水和碳酸氢盐,导致胰液分泌量剧增,此外还能与G细胞相拮抗,抑制胃泌素的释放和胃酸的分泌
十、消化管的血管、淋巴管和神经
(一)血管
供应胃肠的血管多走行于系膜内,动脉穿过肌层,在粘膜下层形成动脉丛。胃和大肠的粘膜下动脉丛发出许多小支进入固有层内,在腺体之间形成毛细血管网,后者汇集成粘膜下静脉丛。静脉伴同动脉穿过肌层离开管壁。小肠的粘膜下动脉丛发出两种分支,一种在肠腺间形成毛细血管网,另一种在绒毛内,紧贴上皮下方形成毛细血管网;两者均汇入粘膜下静脉丛(图12-25)。
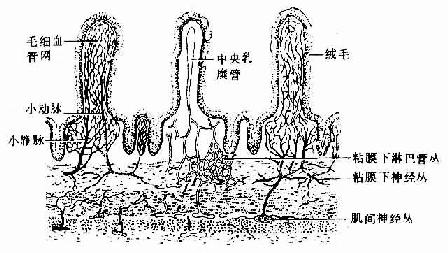
图12-25 消化管的血管、淋巴管与神经分布模式图
(二)淋巴管
胃肠粘膜固有层的腺体间有毛细淋巴管网,淋巴小结周围也有密集的淋巴管网,小肠绒毛内还有中央乳糜管。它们均汇合穿过粘膜肌层,进入粘膜下层形成淋巴管丛。淋巴管伴同血管穿过肌层离开管壁(图12-25)。
(三)神经
支配食管与胃肠的神经由三部分构成:①主要来源于交感神经节与腹腔神经节的节后纤维构成的交感神经;②迷走神经的节前纤维;③由肌间神经丛和粘膜下神经丛内的107~108个神经元及其神经纤维构成的“肠神经系统”(enteric nervce system)。交感神经与迷走神经主要伴随系膜血管进入消化管。前者直接终止于肌层与血管,后者则与肠神经系统中的运动神经元形成突触。切除交感和迷走神经后,动物的消化功能仍能进行,表明肠神经系统具有高度自主性。神经丛由神经节与无髓神经纤维构成,每个神经节含3~50个或更多的神经元。这些神经元可大致分为两型,即中间神经元与运动神经元。后者的轴突止于肌层平滑肌、粘膜肌或腺体。按所含神经递质进行分类,则有胆碱能神经元,5-羟色胺能神经元、肽能神经元等。所有神经元借神经纤维相互连接为功能统一的网络(图12-22)

 中医世家
中医世家 浦 标 网
浦 标 网 河南大学精品课程
河南大学精品课程 图书资料室
图书资料室