脱氧核糖核酸(DNA)是生物界遗传的主要物质基础。生物有机体的遗传特征以密码(code)的形式编码在DNA分子上,表现为特定的核苷酸排列顺序-即遗传信息,在细胞分裂前通过DNA的复制(Replication),将遗传信息由亲代传递给子代,在后代的个体发育过程中,遗传信息自DNA转录(Transcription)给RNA,并指导蛋白质合成,以执行各种生命功能,使后代表现出与亲代相似的遗传性状,这种遗传信息的传递方向,是从DNA到RNA再到蛋白质,即所谓的生物学“中心法则”,80年代以后在某些致癌RNA病毒中发现遗传信息也可存在于RNA分子中,由RNA通过逆转录(reversetranscription)的方式将遗传信息传递给DNA。这为中心法则加入了新的内容。目前认为生物界遗传信息传递的中心法则为:
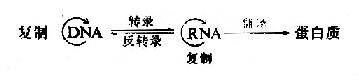
本章的内容主要涉及DNA生物合成的三个方面,第一,DNA复制,第二,RNA反转录为DNA,第三,细胞内DNA受到损伤时进行的修复作用。
第一节 DNA的复制
DNA做为遗传物质的基本特点就是在细胞分裂前进行准确地自我复制(self?replication),使DNA的量成倍增加,这是细胞分裂的物质基础。1953年Watson和Crick提出DNA双螺旋结构模型指出,DNA是由二条互补的脱氧核苷酸链组成,所以一条DNA链上的核苷酸排列顺序是由图16-1双螺旋DNA的复制另一条决定的。这就说明DNA的复制是由原来存在的分子为模板来合成新的链。曾经有过多种关于DNA复制方式的学说,包括半保留复制,全保留复制以及分散复制等(图16-1)。
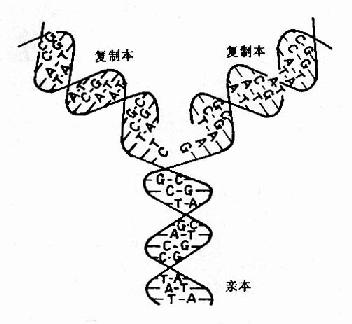
图16-1 双螺旋DNA的复制
一、DNA复制的方式及一般过程:
(一)DNA的半保留复制(semiconservative replication)
Watson和Crick在提出DNA双螺旋结构模型时即推测,DNA在复制时首先两条链之间的氢键断裂两条链分开,然后以每一条链分别做模板各自合成一条新的DNA链,这样新合成的子代DNA分子中一条链来自亲代DNA,另一条链是新合成的,这种复制方式为半保留复制。
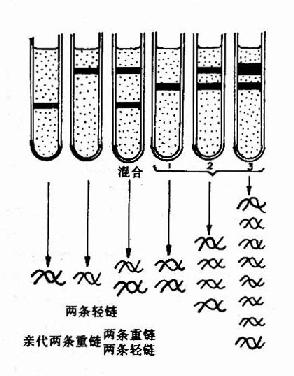
1958年Meselson和Stahl利用氮标记技术在大肠杆菌中首次证实了DNA的半保留复制,他们将大肠杆菌放在含有15N标记的NH4Cl培养基中繁殖了15代,使所有的大肠杆菌DNA被15N所标记,可以得到15N桪NA。然后将细菌转移到含有14N标记的NH4Cl培养基中进行培养,在培养不同代数时,收集细菌,裂介细胞,用氯化铯(CsCl)密度梯度离心法观察DNA所处的位置。由于15N桪NA的密度比普通DNA(14N-DNA)的密度大,在氯化铯密度梯度离心(density gradientcentrifugation)时,两种密度不同的DNA分布在不同的区带。
实验结果表明:在全部由15N标记的培养基中得到的15N桪NA显示为一条重密度带位于离心管的管底。当转入14N标记的培养基中繁殖后第一代,得到了一条中密度带,这是15N桪NA和14N-DNA的杂交分子。第二代有中密度带及低密度带两个区带,这表明它们分别为15N14N-DNA和14N14N-DNA。随着以后在14N培养基中培养代数的增加,低密度带增强,而中密度带逐渐减弱,离心结束后,从管底到管口,CsCl溶液密度分布从高到低形成密度梯度,不同重量的DNA分子就停留在与其相当的CsCl密度处,在紫外光下可以看到DNA分子形成的区带。为了证实第一代杂交分子确实是一半15N-DNA-半14N-DNA,将这种杂交分子经加热变性,对于变性前后的DNA分别进行CsCl密度梯度离心,结果变性前的杂交分子为一条中密度带,变性后则分为两条区带,即重密度带(15N-DNA)及低密度带(14N-DNA)。它们的实验只有用半保留复制的理论才能得到圆满的解释(图16-2和16-3)。
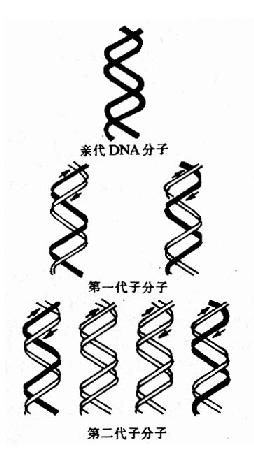 |
 |
| 图16-2 DNA的半保留复制第一代分子含有一条亲代的链(用黑色素示),与另一条新合成的链(用白色表示)配对。在以后的连续复制过程中,原来亲代的两条链仍然保持完整,因此总有两个分子各具有一条原来亲代的链。 |
图16-3 DNA的半保留复制-Meslson?Stahl实验密度梯度离心后的DNA位置:左三管为对照;右三管为实验结果 |
(二)DNA复制的一般过程:
DNA双螺旋是由两条方向相反的单链组成,复制开始时,双链打开,形成一个复制叉(replicative fork,从打开的起点向一个方向形成)或一个复制泡(replicative bubble,从打开的起点向两个方向形成。)两条单链分别做模板。各自合成一条新的DNA链。由于DNA一条链的走向是5′→3′方向,另一条链的走向是3′→5′方向,但生物体内DNA聚合酶只能催化DNA从5′→3′的方向合成。那么,两条方向不同的链怎样才能做模板呢?这个问题由日本学者岗崎先生解决。
原来,在以3′→5′方向的母链为模板时,复制合成出一条5′→3′方向的前导链(leadingstrand),前导链的前进方向与复制叉打开方向是一致的,因此前导链的合成是连续进行的,而另一条母链DNA是5′→3′方向,它作为模板时,复制合成许多条5′→3′方向的短链,叫做随从链(lagging strand),随从链的前进方向是与复制叉的打开方向相反的。随从链只能先以片段的形式合成,这些片段就叫做岗崎片段(Okazaki fragments),原核生物岗崎片段含有1000-2000核苷酸,真核生物一般100?00核苷酸。最后再将多个岗崎片段连接成一条完整的链。由于前导链的合成是连续进行的,而随从链的合成是不连续进行的,所以从总体上看DNA的复制是半不连续复制(图16-4)。
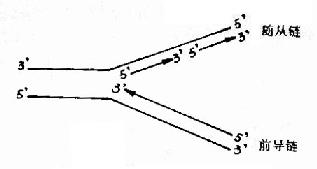
图16-4 DNA的半不连续复制
DNA复制的全部过程可以人为地分成三个阶段,第一个阶段为DNA复制的起始阶段,这个阶段包括起始点,复制方向以及引发体的形成,第二阶段为DNA链的延长,包括前导链及随从链的形成和切除RNA引物后填补空缺及连接岗崎片段。第三阶段为DNA复制的终止阶段。在DNA复制的整个过程中需要30多种酶及蛋白质分子参加,我们将在DNA复制的各个阶段中着重介绍它们的作用。
二、DNA复制的起始阶段:
(一)DNA复制的起始点
很多实验都证明:复制是从DNA分子上的特定部位开始的,这一部位叫做复制起始点(originof replication)常用ori或o表示。细胞中的DNA复制一经开始就会连续复制下去,直至完成细胞中全部基因组DNA的复制。DNA复制从起始点开始直到终点为止,每个这样的DNA单位称为复制子或复制单元(replicon)。在原核细胞中,每个DNA分子只有一个复制起始点,因而只有一个复制子,而在真核生物中,DNA的复制是从许多起始点同时开始的,所以每个DNA分子上有许多个复制子。
DNA复制起始点有结构上的特殊性,例如:大肠杆菌染色体DNA复制起始点Oric由422个核苷酸组成,是一系列对称排列的反向重复序列,即回文结构(palindrome),其中有9个核苷酸或13个核苷酸组成的保守序列,这些部位是大肠杆菌中DnaA蛋白识别的位置,大肠杆菌染色体DNA是环状双链DNA,它的复制是典型的“θ”型复制(由于形状像希腊字母θ)。从一个起点开始,同时向两个方向进行复制,当两个复制方向相遇时,复制就停止。而有些生物的DNA复制起始区是一段富含A·T的区段。这些特殊的结构对于在DNA复制起始过程中参与的酶和许多蛋白质分子的识别和结合都是必须的。
(二)DNA复制的方向:
(1)定点开始双向复制:
这是原核生物和真核生物DNA复制最主要的形式,从一个特定位点解链,沿着两个相反的方向各生长出两条链,形成一个复制泡,用电子显微镜可以观察到复制泡的存在(图16-5)。
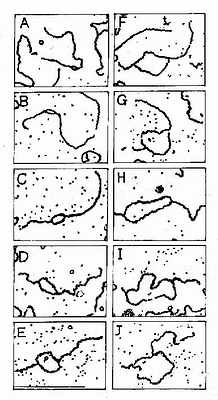
图16-5 SV40DNA;复制泡生长的电镜图谱
(2)定点开始单向复制:
质粒colE1是个典型的例子,复制从一个起始点开始,以同一方向生长出两条链,形成一个复制叉(replication fork)。
(3)两点开始单向复制:
腺病毒DNA的复制是从两个起点开始的,形成两个复制叉,各以一个单一方向复制出一条新链(图16-6)。
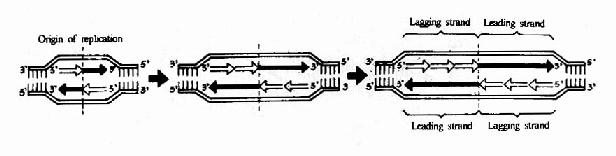
图16-6 DNA的半不连续复制和复制泡的形成
总之DNA复制的起点及方向不仅原核细胞与真核细胞不同,就是同属于原核生物和真核生物的不同种属也有相当大的差异(图16-7)。
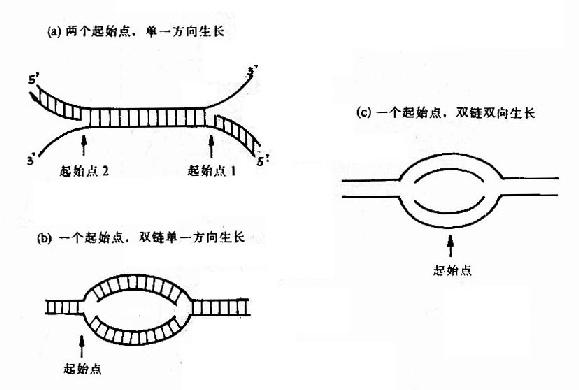
图16-7 DNA链生长方向的三种机制
(三)DNA复制起始引发体的形成及所参与的酶和蛋白质:
1.解链酶(helicase)
DNA开始复制时首先在起始点处解开双链,反应是在一种解链酶(helicase)的催化下进行的。解链酶需要ATP分解供给能量。大肠杆菌中DnaB蛋白就有介链酶活性,与随从链的模板DNA结合,沿5′→3′方向移动,还有一种叫做Rep蛋白和前导链的模板DNA结合沿3′→5′方向移动。解链酶的作用就是打开DNA双链之间的氢键。
2.单链结合蛋白:(single strand binding proteins,SSBP)
它与解开的单链DNA结合,使其稳定不会再度螺旋化并且避免核酸内切酶对单链DNA的水解,保证了单链DNA做为模板时的伸展状态,SSBP可以重复利用(图16-8)。
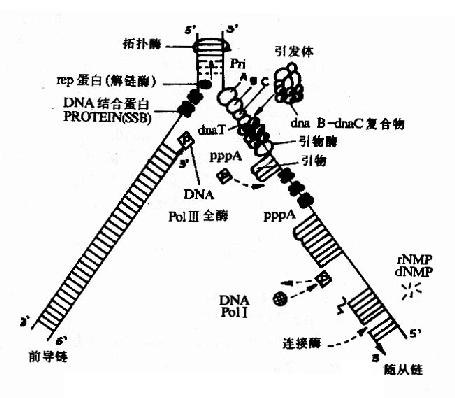
图16-8 大肠杆菌DNA复制叉中复制过程简图
3.引发体的形成:
DNA复制起始的关健步骤是前导链DNA的合成,一旦前导链DNA的聚合作用开始,随从链DNA的合成也随着开始。由于前导链的合成是连续进行的,所以它的起始相对简单,而随从链的合成是不连续进行的,所以引发阶段比较复杂。大肠杆菌的引发前体由Dna B. Dna C和单链结合蛋白组成。
(1)引物酶(primase)
它是一种特殊的RNA聚合酶,可催化短片段RNA的合成。这种短RNA片段一般十几个至数十个核苷酸不等,它们在DNA复制起始处做为引物。RNA引物的3′桹H末端提供了由DNA聚合酶催化形成DNA分子第一个磷酸二酯键的位置。
(2)引发体(primosome)
高度解链的模板DNA与多种蛋白质因子形成的引发前体促进引物酶结合上来,共同形成引发体,引发体主要在DNA随从链上开始,它连续地与引物酶结合并解离,从而在不同部位引导引物酶催化合成RNA引物,在引物RNA的3′桹H末端接下去合成DNA片段,这就是随从链不连续合成的开始。
三、DNA复制的延长阶段以及参与的酶和蛋白质分子:
DNA的复制实际上就是以DNA为模板在DNA聚合酶作用下,将游离的四种脱氧单核苷酸(dATP,dGTP,dCTP,dTTP,简写为dNTP)聚合成DNA的过程。
这是一个非常复杂的酶促反应,需要许多种酶和蛋白质参与,现分别叙述它们在DNA复制中作用。
(一)DNA的聚合反应和DNA聚合酶
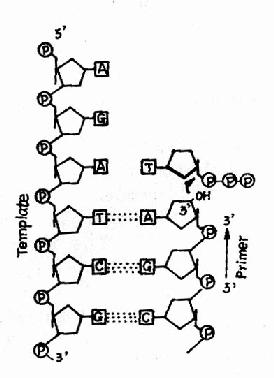
图16-9 DNA聚合酶的作用
1957年,Arthur kornberg首次在大肠杆菌中发现DNA聚合酶Ⅰ,(DNa polymerase Ⅰ,简写DNA polⅠ)后来又相继发现了DNA聚合酶Ⅱ和DNA聚合酶Ⅲ。(DNa polymerase Ⅱ,Ⅲ,简写DNA polⅡ,DNA polⅢ)实验证明大肠杆菌中DNA复制的主要过程靠DNa polⅢ起作用,而DNA polⅠ和DNA polⅡ在DNA错配的校正和修复中起作用。见表16-1。
这种酶的共同性质是:①需要DNA模板,因此这类酶又称为依赖DNA的DNA聚合酶(DNa dependent DNA polymerase, DDDP)。②需要RNA或DNA做为引物(primer),即DNA聚合酶不能从头催化DNA的起始。③催化dNTP加到引物的3′桹H末端,因而DNA合成的方向是5′→3′。图16-9。④三种DNA聚合酶都属于多功能酶,它们在DNA复制和修复过程的不同阶段发挥作用。由于DNA聚合酶Ⅰ是研究得最清楚而且代表了其他DNA聚合酶的基本特点,所以我们着重介绍DNa polⅠ的作用并指出另外二种DNA pol的特殊性:
1.DNA聚合酶Ⅰ:
DNA polⅠ是由一条多肽链组成,分子量为109KD。酶分子中含有一个Zn++,是聚合活性必须的。
大肠杆菌每个细胞中约有400个酶分子,每个酶分子每分钟在37℃下能催化667个核苷酸参入到DNA链中,用枯草杆菌蛋白酶可将此酶水介成两个片段,大片段分子量为76KD,通常称为klenow片段,小片段为34KD。大小片段具有不同的酶活性。
(1)DNA聚合酶的5′→3′聚合活性:
这是DNA聚合酶最主要的活性,按模板DNA上的核苷酸顺序,将互补的dNTP逐个加到引物RNA3′桹H末端,并促进3′桹H与dNTP的5′桺O4形成磷酸二酯键,酶的专一性表现为新进入的dNTP必须与模板DNA碱基配对时才有催化作用,5′→3′聚合活性存在于klenow片段上(图16-9和图16-10)。
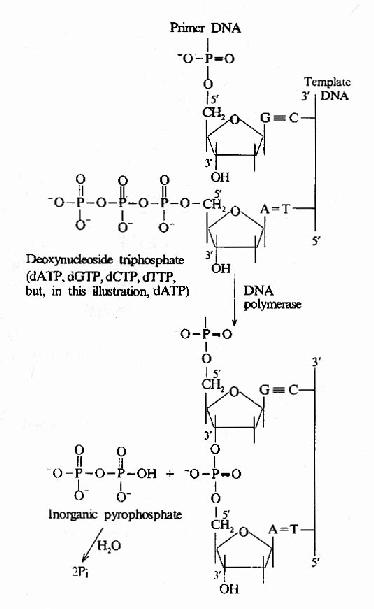
图16-10 DNA聚合酶催化的DNA链延长
(2)DNA聚合酶的3′→5′外切核酸酶活性:
这种酶活性的主要功能是从3′→5′方向识别并切除DNA生长链末端与模板DNA不配对而游离的核苷酸,这种功能称为校对功能,这是保证其聚合作用的正确性不可缺少的,因此对于DNA复制中极高的保真性是至关重要的。
(3)DNA聚合酶的5′→3′外切核酸酶活性:
这种酶活性是从DNA链的5′端向3′末端水解已配对的核苷酸,本质是切断磷酸二酯键,每次能切除10个核苷酸。因此,这种酶活性在DNA损伤的修复中可能起重要作用,对完成的DNA片段去除5′端的RNA引物也是必须的。
DNA polⅠ的5′→3′聚合活性和5′→3′外切酶活性协同作用,可以使DNA一条链上的切口从5′→3′方向移动,这种反应叫做缺刻平移(nick translation),利用此反应可在体外对DNA片段进行放射性磷(α-32PdNTP)的标记制成探针(probe),进行核酸的分子杂交实验,是现代分子生物学的一项重要技术(图16-11)。
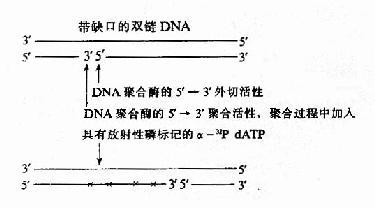
图16-11 缺刻平移标记DNA探针
许多实验证实DNA polⅠ并不是DNA复制过程中的主要酶,它的作用主要与DNA损伤后的修复有关。
2.DNA聚合酶Ⅱ(DNA polⅡ)
此酶分子量为120KD,每个细胞约有100个酶分子,但活性只有DNa polⅠ的5%,它具有5′→3′聚合活性和3′→5′外切活性,而没有5′→3′外切活性,它的作用可能与DNA损伤修复有关。
3.DNA聚合酶Ⅲ(DNA polⅢ)
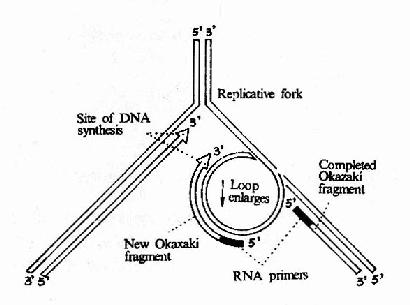
图16-12 DNA聚合酶Ⅲ催化先导链和随从的合成
这是在DNA复制过程中起主要作用的聚合酶,它是由一个多亚基组成的蛋白质分子,其分子量>600kDa整个酶分子形成一个不对称的二聚体,每个大肠杆菌细胞中只有10?0个酶分子,但催化dNTP参入DNA链的速率却是最快的,约为9000核苷酸/每分钟/每个酶分子。这也证明DNa polⅢ是DNA复制过程中主要发挥作用的酶。在大肠杆菌染色体DNA进行复制时,DNA聚合酶Ⅲ全酶并不是单独起作用的,而是与引发体,介链酶等构成一个复制体(replisome)。由于复制体的存在,先导链和随从链可以同时复制。DNa polⅢ是由多亚基组成的不对称二聚体,它可能同时负责先导链和随从链的复制,在φ×174的复制中观察到引发体总是伴随着DNA噜噗(loop)的存在。图16?2可以看到,由于随从链的模板DNA在DNA聚合酶Ⅲ全酶上绕转了180°而形成一个噜噗,因此岗崎片段的合成方向能够与先导链的合成方向以及复制体移动方向保持一致。
随着DNA polⅢ向前移动,先导链的合成逐渐延长的同时,岗崎片段也在不断延长,这一噜噗也在不断扩大。当岗崎片段合成到前一个片段的5′端时,这一大噜噗就释放出来,由于复制叉向前移动又可将另一部分随从链的模板置换出来,由引发体合成新的引物,然后再形成一个小的噜噗,进行新的岗崎片段的合成。由此模型不难看出:随从链的合成需要周期性的引发,因此其合成进度总是与前导链相差一个岗崎片段的长度。岗崎片段完成后,其5′端的RNA引物由DNa polⅠ的5′→3′外切酶活性切除,由此造成的空隙再由DNA polⅠ的5′→3′聚合活性催化dNTP得到填补。所以DNA的复制是在DNa polⅢ和DNApolⅠ互相配合下完成的。
下面列表说明三种大肠杆菌DNA聚合酶的特性
表16-1 大肠杆菌DNA聚合酶特征
|
DNA聚合酶Ⅰ |
DNA聚合酶Ⅱ |
DNA聚合酶Ⅲ |
| 分子量 |
109KD |
120KD |
>600KD |
| 每个细胞中的分子数 |
400 |
17-100 |
10-20 |
| 5′→3′聚合活性 |
+ |
+ |
+ |
| 37℃转化率核苷酸数/酶分子·分钟 |
600 |
30 |
30,000 |
| 5′→3′外切活性 |
+ |
- |
- |
| 5′→3′外切活性 |
+ |
+ |
+ |
| 切刻平移活性 |
+ |
- |
- |
| 对dNTP亲和力 |
低 |
低 |
高 |
| 功能 |
修复 |
不详 |
复制 |
| 去除引物 |
|
|
| 填补空缺 |
|
|
真核生物DNA聚合酶
真核生物DNA聚合酶有α、β、γ、δ及ε。它们的基本特性相似于大肠杆菌DNA聚合酶,其主要活性是催化dNTP的5′→3′聚合活性,基本特征见表16-2。
表16-2 真核生物DNA聚合酶
|
α |
β |
γ |
δ |
ε |
| 亚基数 |
4 |
4 |
4 |
2 |
5 |
| 分子量(KD) |
>250 |
36-38 |
160-300 |
170 |
256 |
| 细胞内定位 |
核 |
核 |
线粒体 |
核 |
核 |
| 5′→3′聚合活性 |
+ |
+ |
+ |
+ |
+ |
| 3′→5′外切活性 |
- |
- |
- |
- |
- |
| 功能 |
复制、引发 |
修复 |
复制 |
复制 |
复制 |
真核细胞在DNA复制中起主要作用的是DNA polα,主要负责染色体DNA的复制。DNa polβ的模板特异性是具有缺口的DNA分子,被认为它与DNA修复有关。DNa polγ在线粒体DNA的复制中起作用。DNA polδ不但有5′→3′聚合活性,而且还具有3′→5′外切酶活性,据认为真核生物DNA复制是在DNa polα和DNA polδ协同作用下进行的,前导链的合成靠DNA polδ催化,并且还需要一种细胞周期调节因子椩鲋诚赴?丝乖?proliferatingcell nucleus antigen, PCNA)参与。而随从链的合成靠DNA polα和引发酶配合作用完成。
(二)与超螺旋松驰有关的酶:
DNA复制从起始点开始向一个方向复制时,局部的DNA双链必须打开,主要靠解链酶的作用,打开后的单链还需要单链结合蛋白与其结合,在复制叉向前移动时造成其前方DNA分子所产生的正超螺旋,必须由拓扑异构酶来解决。下面分别介绍它们的作用。
拓扑异构酶(topoisomerase)是一类改变DNA拓扑性质的酶。在体外可催化DNA的各种拓扑异构化反应,而在生物体内它们可能参与了DNA的复制与转录。在DNA复制时,复制叉行进的前方DNA分子部分产生有正超螺旋,拓扑酶可松驰超螺旋,有利于复制叉的前进及DNA的合成。DNA复制完成后,拓扑酶又可将DNA分子引入超螺旋,使DNA缠绕、折叠,压缩以形成染色质。DNA拓扑异构酶有Ⅰ型和Ⅱ型,它们广泛存在于原核生物及真核生物中。表16-3
表16-3 大肠杆菌和真核生物中的拓扑异构酶
| 类型 |
作用 |
对超螺旋的作用 |
| Ⅰ型拓扑异构酶 |
|
|
| 大肠杆菌 |
切开一股DNA链 |
松驰负超螺旋 |
| 真核生物 |
切开一股DNA链 |
松驰正,负超螺旋 |
| Ⅱ型拓扑异构酶 |
|
|
| 大肠杆菌 |
切开二股DNA链 |
构驰正超螺旋; |
| 依赖ATP |
引入负超螺旋, |
|
解环连等 |
| 真核生物 |
切开二股DNA链 |
松驰正超旋, |
| 依赖ATP |
但不能引入负超螺旋 |
拓扑异构酶Ⅰ(TopoⅠ)的主要作用是将环状双链DNA的一条链切开一个口,切口处链的末端绕螺旋轴按照松驰超螺旋的方向转动,然后再将切口封起来。这就使DNA复制叉移动时所引起的前方DNA正超螺旋得到缓解,利于DNA复制叉继续向前打开。拓扑异构酶Ⅰ除上述作用外,对环状单链DNA还有打结或解结作用,对环状双链DNA的环连或解连以及使环状单链DNA形成环状双链DNA都有作用(图16-13)。
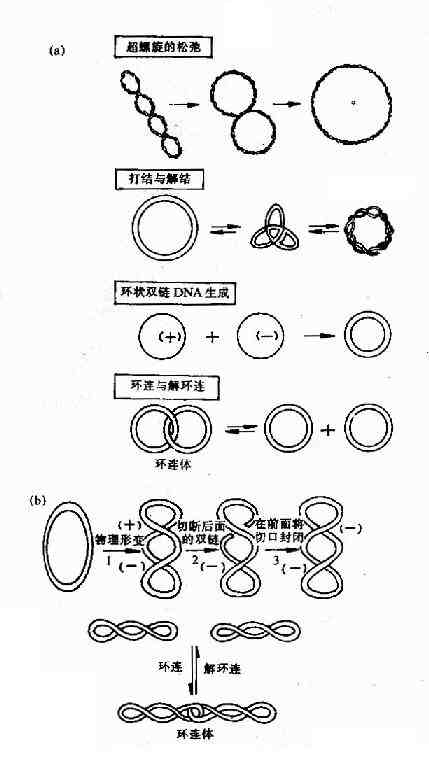
图16-13 拓扑酶Ⅰ及Ⅱ的作用特点
(a)大肠杆菌拓扑酶Ⅰ催化的4种拓扑异构化作用 (b)拓扑酶Ⅱ的作用
拓扑异构酶Ⅱ(TopoⅡ)是在大肠杆菌中发现的,曾被称为旋转酶(gyrase),它们作用特点是切开环状双链DNA的两条链,分子中的部分经切口穿过而旋转,然后封闭切口,TopoⅡ还可使DNA分子从超螺旋状态转变为松驰状态,此反应不需要ATP参与。DNA复制完成后,TopoⅡ在ATP参与下,DNA分子从松驰状态转变为负超螺旋。此外,TopoⅡ催化的拓扑异构化反应还有环连或解环连,以及打结或解结。
四、DNA复制的终止阶段
DNA在复制过程中,合成出的前导链为一条连续的长链。随从链则是由合成出许多相邻的片段,在连接酶的催化下,连接成为一条长链。连接作用是在连接酶催化下进行的。
连接酶(ligase)的作用是催化相邻的DNA片段以3′、5′-磷酸二酯键相连接。连接反应中的能量来自ATP(或NAD+)。连接酶先与ATP作用,以共价键相连生成E桝MP中间体。中间体即与一个DNA片段的5′-磷酸相连接形成E-AMP-5′-DNA。然后再与另一个DNA片段的3′-OHH末端作用,E和AMP脱下,两个DNA片段以3′、5′磷酸二酯键相连接。随从链的各个DNA片段就是这样连接成一条DNA长链(图16-14)。?
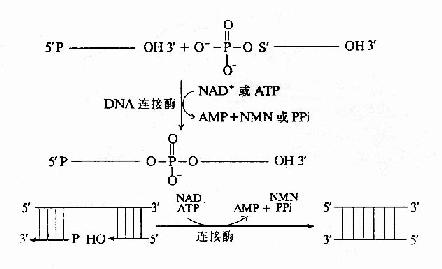 |
 |
| 图16-14 连接酶的催化反应 |
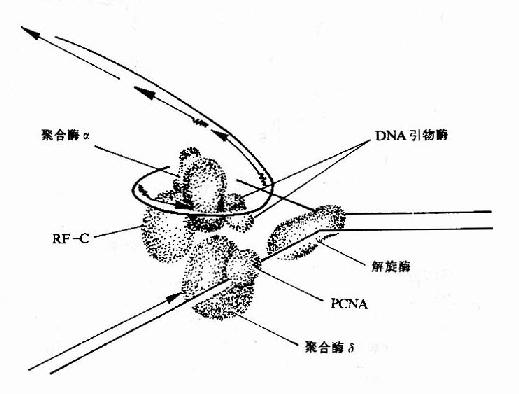
图16-15 真核生物DNA复制叉结构示意图 |
已有研究证明大肠杆菌染色体DNA具有复制终止位点,此处可以结合一种特异的蛋白质分子叫做Tus,这个蛋白质可能是通过阻止解链酶(Helicase)的解链活性而终止复制的。详细的机制还不完全清楚。
DNA复制完成后,靠拓扑酶将DNA分子引入超螺旋结构。
五、真核生物DNA复制的特点:
DNA复制的研究最初是在原核生物中进行的,有些原核生物的DNA复制已经搞得很清楚。真核生物比原核生物复杂得多,但DNA复制的基本过程还是相似的。在这里我们主要讨论一些重要的区别。
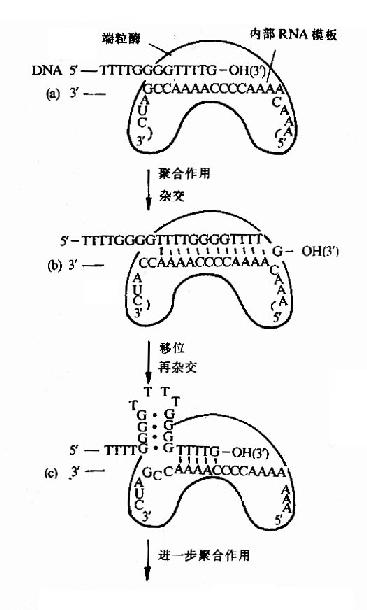
图16-16 端粒酶催化端区TG链的合成
1.与原核生物不同,真核生物DNA复制有许多起始点,例如酵母S.cerevisiae的17号染色体约有400个起始点,因此,虽然真核生物DNA复制的速度(60核苷酸/每秒钟)比原核生物DNA复制的速度(E.coli1700核苷酸/每秒钟)慢得多,但复制完全部基因组DNA也只要几分钟的时间。
2.SV40病毒DNA主要依靠宿主细胞中的DNA复制体系进行DNA的复制,这是了解真核生物DNA复制的体外模型。在真核生物DNA复制叉处,需要两种不同的酶。DNA聚合酶α(polα)和DNA聚合酶δ(polδ)。polα和引物酶紧密结合,在DNA模板上先合成RNA引物,再由polα延长DNA链,这种活性还要复制因子C参与。同时结合在引物模板上的PCNA(增殖细胞核抗原Proliferating cell nuclear antigen)此时释放了polα,然后由polδ结合到生长链3′末端,并与PCNA结合,继续合成前导链。而随从链的合成靠polα紧密与引物酶结合并在复制因子C帮助下,合成岗崎片段(图16-15)。
3.由于真核生物染色体是线性DNA,它的两端叫做端区(telomeres),端区是由重复的寡核苷酸序列构成的。例如酵母的端区重复序列是5′G(1?)T(3)3′。前面讲到所有生物DNA聚合酶都只能催化DNA从5′→3′的方向合成,因此当复制叉到达线性染色体末端时,前导链可以连续合成到头,而由于随从链是以一种不连续的形式合成岗崎片段,所以不能完成线性染色体末端的复制,如果这个问题不解决,真核生物在细胞分裂时DNA复制将产生5′末端隐缩,使DNA缩短,近十多年的研究表明,真核生物体内都存在一种特殊的反转录酶叫做端粒酶(telomerase),它是由蛋白质和RNA两部分组成的,它以自身的RNA为模板,在随从链模板DNA的3′桹H末端延长DNA,再以这种延长的DNA为模板,继续合成随从链(图16-16)。
由此可见端粒酶在保证染色体复制的完整性上有重要意义。
第二节 反转录作用(reverse transcription)
1970年Temin等在致癌RNA病毒中发现了一种特殊的DNA聚合酶,该酶以RNA为核板,根据碱基配对原则,按照RNA的核苷酸顺序(其中V与A配对)合成DNA。这一过程与一般遗传信息流转录的方向相反,故称为反转录,催化此过程的DNA聚合酶叫做反转录酶(reverse transcriptase)。后来发现反转录酶不仅普遍存在于RNA病毒中,哺乳动物的胚胎细胞和正在分裂的淋巴细胞中也有反转录酶。
反转录酶的作用是以dNTP为底物,以RNA为模板,tRNA(主要是色氨酸tRNA)为引物,在tRNA3′桹H末端上,按5′→3′方向,合成一条与RNA模板互补的DNA单链,这条DNA单链叫做互补DNA(complementary DNA, cDNA),它与RNA模板形成RNA桪NA杂交体。随后又在反转录酶的作用下,水解掉RNA链,再以cDNA为模板合成第二条DNA链。至此,完成由RNA指导的DNA合成过程(图16-17)。
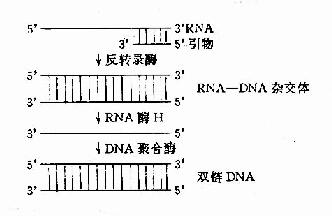
图16-17 反转录酶催化的反转录作用
携带反转录酶的病毒又称为反转录病毒,它侵入宿主细胞后先以病毒RNA为模板靠反转录酶催化合成DNA,随后这种DNA环化并整合到宿主细胞的染色体DNA中去,以原病毒(provirus)的形式在宿主细胞中一代代传递下去。以后又发现许多反转录病毒基因组中都含有癌基因(oncogene),如果由于某种因素激活了癌基因就可使宿主细胞转化为癌细胞。(将在专门一章中叙述)。
大多数反转录酶都具有多种酶活性,主要包括以下几种活性。①DNA聚合酶活性;以RNA为模板,催化dNTP聚合成DNA的过程。此酶需要RNA为引物,多为色氨酸的tRNA,在引物tRNA3′-末端以5′→3′方向合成DNA。反转录酶中不具有3′→5′外切酶活性,因此没有校正功能,所以由反转录酶催化合成的DNA出错率比较高。②RNase H活性;由反转录酶催化合成的cDNA与模板RNA形成的杂交分子,将由RNaseH从RNA5′端水解掉RNA分子。③DNA指导的DNA聚合酶活性;以反转录合成的第一条DNA单链为模板,以dNTP为底物,再合成第二条DNA分子。除此之外,有些反转录酶还有DNA内切酶活性,这可能与病毒基因整合到宿主细胞染色体DNA中有关。反转录酶的发现对于遗传工程技术起了很大的推动作用,目前它已成为一种重要的工具酶。用组织细胞提取mRNA并以它为模板,在反转录酶的作用下,合成出互补的DNA(cDNA),由此可构建出cDNA文库(cDNa library),从中筛选特异的目的基因,这是在基因工程技术中最常用的获得目的基因的方法,详见基因工程一章。
小结
DNA是携带遗传信息的载体,细胞分裂前,通过DNA的复制作用,遗传信息从亲代DNA分子传到子代DNA分子中。DNA复制时,是从一个特定的起始点开始的,两条DNA链分别做模板,在DNA聚合酶等许多酶和蛋白质分子的参与下,以四种脱氧单核苷酸为原料(dNTP),以碱基配对为原则合成新一代的DNA分子,合成方向是5′→3′。经过复制后的DNA分子中,一条链来自亲代DNA分子,另一条链是新合成的,这种复制方式叫做半保留复制。由于DNA分子复制时,两条链分别做模板,有一条链是连续合成的,这条链为前导链,而另一条链合成时只能以5′→3′方向先合成岗崎片段,然后再靠连接酶将这些片段连接起来,形成随从链,所以DNA复制是半不连续的合成。
DNA复制时,先要由DNA拓扑异构酶作用于DNA分子双螺旋,使之松驰,然后才能由解链酶作用,解开双链,此时引物酶合成一段RNA分子做为引物,在DNA聚合酶Ⅲ催化下,连续地合成DNA链。随从链的合成靠多种酶和蛋白质因子参与,在引物酶作用下合成RNA引物,在DNA聚合酶Ⅲ作用下合成DNA片段,它们共同形成了岗崎片段,RNA引物是靠DNA聚合酶Ⅰ的5'→3'外切活性切除的,切除引物后的空隙,再靠DNA聚合酶Ⅰ填补,最后在连接酶作用下,形成长链。
真核生物中DNA的复制靠DNA聚合酶,它有多种不同的形式(α、β、γ、δ等)此外,还有像PCNA等多种蛋白质因子参与。
DNA复制中的准确性很高,因为原核细胞靠DNA聚合酶Ⅰ而真核细胞靠DNA聚合酶δ,都具有3′→5′外切酶活性,可以校正复制中出现的碱基错配。DNA的合成也可以靠反转录酶的催化而完成,这是以RNA分子为模板,合成DNA分子的过程,在致癌的RNA病毒中,有反转录酶的存在,真核生物中的端粒酶就是一种反转录酶,它催化染色体端区DNA的合成。
思考题
1.DNA复制起始有哪些特点?
2.原核生物与真核生物中的DNA聚合酶有哪些种类,它们各自的功能是什么?
3.DNA复制时前导链与随从链的合成有哪些不同?
4.DNA拓扑异构酶的作用如何?
5.连接酶催化的连接反应与DNA聚合酶5′→3′聚合活性有何异同?
6.什么是反转录,有什么生物学意义?
第三节 DNA的损伤与修复
DNA存储着生物体赖以生存和繁衍的遗传信息,因此维护DNA分子的完整性对细胞至关重要。外界环境和生物体内部的因素都经常会导致DNA分子的损伤或改变,而且与RNA及蛋白质可以在胞内大量合成不同,一般在一个原核细胞中只有一份DNA,在真核二倍体细胞中相同的DNA也只有一对,如果DNA的损伤或遗传信息的改变不能更正,对体细胞就可能影响其功能或生存,对生殖细胞则可能影响到后代。所以在进化过程中生物细胞所获得的修复DNA损伤的能力就显得十分重要,也是生物能保持遗传稳定性之所在。在细胞中能进行修复的生物大分子也就只有DNA,反映了DNA对生命的重要性。另一方面,在生物进化中突变又是与遗传相对立统一而普遍存在的现象,DNA分子的变化并不是全部都能被修复成原样的,正因为如此生物才会有变异、有进化。
一、DNA的损伤
(一)DNA损伤的原因
1.DNA分子的自发性损伤
(1)DNA复制中的错误 以DNA为模板按碱基配对进行DNA复制是一个严格而精确的事件,但也不是完全不发生错误的。碱基配对的错误频率约为10-1-10-2,在DNA复制酶的作用下碱基错误配对频率降到约10-5-10-6,复制过程中如有错误的核苷酸参入,DNA聚合酶还会暂停催化作用,以其3′-5′外切核酸酶的活性切除错误接上的核苷酸,然后再继续正确的复制,这种校正作用广泛存在于原核和真核的DNA聚合酶中,可以说是对DNA复制错误的修复形式,从而保证了复制的准确性。但校正后的错配率仍约在10-10左右,即每复制1010个核苷酸大概会有一个碱基的错误。
(2)DNA的自发性化学变化 生物体内DNA分子可以由于各种原因发生变化,至少有以下类型:
a.碱基的异构互变 DNA中的4种碱基各自的异构体间都可以自发地相互变化(例如烯醇式与酮式碱基间的互变),这种变化就会使碱基配对间的氢键改变,可使腺嘌呤能配上胞嘧啶、胸腺嘧啶能配上鸟嘌呤等,如果这些配对发生在DNA复制时,就会造成子代DNA序列与亲代DNA不同的错误性损伤,如图16-18所示。
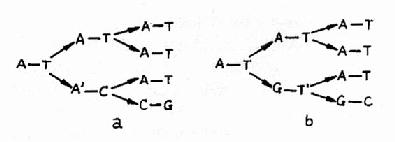
图16-18 腺嘌呤的稀有互变异体与胞嘧啶
(a),或胸腺嘧啶的稀有互变异构体与鸟嘌呤
(b)的氢链形成导致下一世代中G-C配对取代A-T配对
b.碱基的脱氨基作用 碱基的环外氨基有时会自发脱落,从而胞嘧啶会变成尿嘧啶、腺嘌呤会变成次黄嘌呤(H)、鸟嘌呤会变成黄嘌呤(X)等,遇到复制时,U与A配对、H和X都与C配对就会导致子代DNA序列的错误变化。胞嘧啶自发脱氨基的频率约为每个细胞每天190个。
c.脱嘌呤与脱嘧啶 自发的水解可使嘌呤和嘧啶从DNA链的核糖磷酸骨架上脱落下来。一个哺乳类细胞在37℃条件下,20h内DNA链上自发脱落的嘌呤约1000个、嘧啶约500个:估计一个长寿命不复制繁殖的哺乳类细胞(如神经细胞)在整个生活期间自发脱嘌呤数约为108,约占细胞DNA中总嘌呤数的3%。
d.碱基修饰与链断裂 细胞呼吸的副产物O2、H2O2等会造成DNA损伤,能产生胸腺嘧啶乙二醇、羟甲基尿嘧啶等碱基修饰物,还可能引起DNA单链断裂等损伤,每个哺乳类细胞每天DNA单链断裂发生的频率约为5万次。此外,体内还可以发生DNA的甲基化,结构的其他变化等,这些损伤的积累可能导致老化。
由此可见,如果细胞不具备高效率的修复系统,生物的突变率将大大提高。
2.物理因素引起的DNA损伤 射线引起的DNA损伤是最引人注意的。
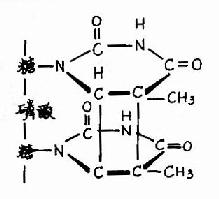
图16-19 胸腺嘧啶二聚体的形成
(1)紫外线引起的DNA损伤 DNA分子损伤最早就是从研究紫外线的效应开始的。当DNA受到最易被其吸收波长(~260nm)的紫外线照射时,主要是使同一条DNA链上相邻的嘧啶以共价键连成二聚体,相邻的两个T、或两个C、或C与T间都可以环丁基环(cyclobutane ring)连成二聚体,其中最容易形成的是TT二聚体,如图16-19所示。
人皮肤因受紫外线照射而形成二聚体的频率可达每小时5×104/细胞,但只局限在皮肤中,因为紫外线不能穿透皮肤。但微生物受紫外线照射后,就会影响其生存。紫外线照射还能引起DNA链断裂等损伤。
(2)电离辐射引起的DNA损伤 电离辐射损伤DNA有直接和间接的效应,直接效应是DNA直接吸收射线能量而遭损伤,间接效应是指DNA周围其他分子(主要是水分子)吸收射线能量产生具有很高反应活性的自由基进而损伤DNA。电离辐射可导致DNA分子的多种变化:
a.碱基变化 主要是由OH-自由基引起,包括DNA链上的碱基氧化修饰、过氧化物的形成、碱基环的破坏和脱落等。一般嘧啶比嘌呤更敏感。
b.脱氧核糖变化 脱氧核糖上的每个碳原子和羟基上的氢都能与OH-反应,导致脱氧核糖分解,最后会引起DNA链断裂。
c.DNA链断裂 这是电离辐射引起的严重损伤事件,断链数随照射剂量而增加。射线的直接和间接作用都可能使脱氧核糖破坏或磷酸二酯键断开而致DNA链断裂。DNA双链中一条链断裂称单链断裂(single strand broken),DNA双链在同一处或相近处断裂称为双链断裂(doublestrand broken)。虽然单链断裂发生频率为双链断裂的10-20倍,但还比较容易修复;对单倍体细胞来说(如细菌)一次双链断裂就是致死事件。
d.交联 包括DNA链交联和DNA-蛋白质交联。同一条DNA链上或两条DNA链上的碱基间可以共价键结合,DNA与蛋白质之间也会以共价键相连,组蛋白、染色质中的非组蛋白、调控蛋白、与复制和转录有关的酶都会与DNA共价键连接。这些交联是细胞受电离辐射后在显微镜下看到的染色体畸变的分子基础,会影响细胞的功能和DNA复制。
3.化学因素引起的DNA损伤
化学因素对DNA损伤的认识最早来自对化学武器杀伤力的研究,以后对癌症化疗、化学致癌作用的研究使人们更重视突变剂或致癌剂对DNA的作用。
(1)烷化剂对DNA的损伤 烷化剂是一类亲电子的化合物,很容易与生物体中大分子的亲核位点起反应。烷化剂的作用可使DNA发生各种类型的损伤:
a.碱基烷基化。烷化剂很容易将烷基加到DNA链中嘌呤或嘧啶的N或O上,其中鸟嘌呤的N7和腺嘌呤的N3最容易受攻击,烷基化的嘌呤碱基配对会发生变化,例如鸟嘌呤N7被烷化后就不再与胞嘧啶配对,而改与胸腺嘧啶配对,结果会使G-C转变成A-T。
b.碱基脱落。烷化鸟嘌呤的糖苷键不稳定,容易脱落形成DNA上无碱基的位点,复制时可以插入任何核苷酸,造成序列的改变。
c.断链。DNA链的磷酸二酯键上的氧也容易被烷化,结果形成不稳定的磷酸三酯键,易在糖与磷酸间发生水解,使DNA链断裂。
d.交联。烷化剂有两类,一类是单功能基烷化剂,如甲基甲烷碘酸,只能使一个位点烷基化;另一类是以双功能基烷化剂,化学武器如氮芥、硫芥等,一些抗癌药物如环磷酰胺、苯丁酸氮芥、丝裂霉素等,某些致癌物如二乙基亚硝胺等均属此类,其两个功能基可同时使两处烷基化,结果就能造成DNA链内、DNA链间,以及DNA与蛋白质间的交联。
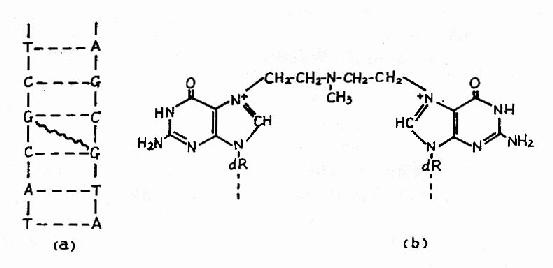
图16-20 氮芥引起DNA分子两条链在鸟嘌呤上的交联
(a)交联附近的总图;(b)交联部分结构图
(2)碱基类似物、修饰剂对DNA的损伤 人工可以合成一些碱基类似物用作促突变剂或抗癌药物,如5-溴尿嘧啶(5-BU)、5-氟尿嘧啶(5-FU)、2-氨基腺嘌呤(2-AP)等。由于其结构与正常的碱基相似,进入细胞能替代正常的碱基参入到DNA链中而干扰DNA复制合成,例如5-BU结构与胸腺嘧啶十分相近,在酮式结构时与A配对,却又更容易成为烯醇式结构与G配对,在DNA复制时导致A-T转换为G-C。
还有一些人工合成或环境中存在的化学物质能专一修饰DNA链上的碱基或通过影响DNA复制而改变碱基序列,例如亚硝酸盐能使C脱氨变成U,经过复制就可使DNA上的G桟变成A桾对;羟胺能使T变成C,结果是A桾改成C桮对;黄曲霉素B也能专一攻击DNA上的碱基导致序列的变化,这些都是诱发突变的化学物质或致癌剂。
(二)DNA损伤的后果
上述损伤会最终导致DNA分子结构的变化,这种DNA分子水平上的突变(mutation)是整体遗传突变的基础。
归纳DNA损伤后分子最终的改变,有以下几种类型:
1.点突变(point mutation) 指DNA上单一碱基的变异。嘌呤替代嘌呤(A与G之间的相互替代)、嘧啶替代嘧啶(C与T之间的替代)称为转换(transition);嘌呤变嘧啶或嘧啶变嘌呤则称为颠换(transvertion)。
2.缺失(deletion) 指DNA链上一个或一段核苷酸的消失。
3.插入(insertion) 指一个或一段核苷酸插入到DNA链中。在为蛋白质编码的序列中如缺失及插入的核苷酸数不是3的整倍数,则发生读框移动(reading frame shift),使其后所译读的氨基酸序列全部混乱,称为移码突变(frame?shift mutaion)。
4.倒位或转位(transposition) 指DNA链重组使其中一段核苷酸链方向倒置、或从一处迁移到另一处。
5.双链断裂 已如前述,对单倍体细胞一个双链断裂就是致死性事件。
突变或诱变对生物可能产生4种后果:①致死性;②丧失某些功能;③改变基因型(genotype)而不改变表现型(phenotype);④发生了有利于物种生存的结果,使生物进化。
二、DNA修复
DNA修复(DNA repairing)是细胞对DNA受损伤后的一种反应,这种反应可能使DNA结构恢复原样,重新能执行它原来的功能;但有时并非能完全消除DNA的损伤,只是使细胞能够耐受这种DNA的损伤而能继续生存。也许这未能完全修复而存留下来的损伤会在适合的条件下显示出来(如细胞的癌变等),但如果细胞不具备这修复功能,就无法对付经常发生的DNA损伤事件,就不能生存。所以研究DNA修复也是探索生命的一个重要方面,而且与军事医学、肿瘤学等密切相关。对不同的DNA损伤,细胞可以有不同的修复反应。
(一)回复修复
这是较简单的修复方式,一般都能将DNA修复到原样。
1.光修复 这是最早发现的DNA修复方式。修复是由细菌中的DNA光解酶(photolyase)完成,此酶能特异性识别紫外线造成的核酸链上相邻嘧啶共价结合的二聚体,并与其结合,这步反应不需要光;结合后如受300-600nm波长的光照射,则此酶就被激活,将二聚体分解为两个正常的嘧啶单体,然后酶从DNA链上释放,DNA恢复正常结构。后来发现类似的修复酶广泛存在于动植物中,人体细胞中也有发现。
2.单链断裂的重接 DNA单链断裂是常见的损伤,其中一部分可仅由DNA连接酶(ligase)参与而完全修复。此酶在各类生物各种细胞中都普遍存在,修复反应容易进行。但双链断裂几乎不能修复。
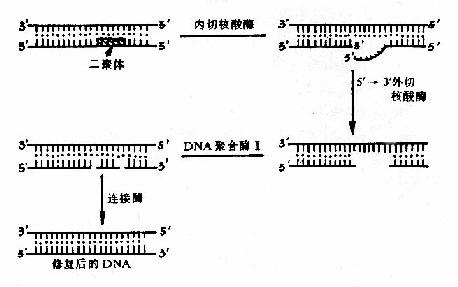
图16-21 损伤DNA的切除修复
3.碱基的直接插入 DNA链上嘌呤的脱落造成无嘌呤位点,能被DNA嘌呤插入酶(insertase)识别结合,在K+存在的条件下,催化游离嘌呤或脱氧嘌呤核苷插入生成糖苷键,且催化插入的碱基有高度专一性、与另一条链上的碱基严格配对,使DNA完全恢复。
4.烷基的转移 在细胞中发现有一种O6甲基鸟嘌呤甲基转移酶,能直接将甲基从DNA链鸟嘌呤O6位上的甲基移到蛋白质的半胱氨酸残基上而修复损伤的DNA。这个酶的修复能力并不很强,但在低剂量烷化剂作用下能诱导出此酶的修复活性。
(二)切除修复(excision repair) 是修复DNA损伤最为普遍的方式,对多种DNA损伤包括碱基脱落形成的无碱基位点、嘧啶二聚体、碱基烷基化、单链断裂等都能起修复作用。这种修复方式普遍存在于各种生物细胞中,也是人体细胞主要的DNA修复机制。修复过程需要多种酶的一系列作用,基本步骤如图16?1所示,①首先由核酸酶识别DNA的损伤位点,在损伤部位的5′侧切开磷酸二酯键。不同的DNA损伤需要不同的特殊核酸内切酶来识别和切割。②由5′→3′核酸外切酶将有损伤的DNA片段切除。③在DNA聚合酶的催化下,以完整的互补链为模板,按5′→3′方向DNA链,填补已切除的空隙。④由DNA连接酶将新合成的DNA片段与原来的DNA断链连接起来。这样完成的修复能使DNA恢复原来的结构。
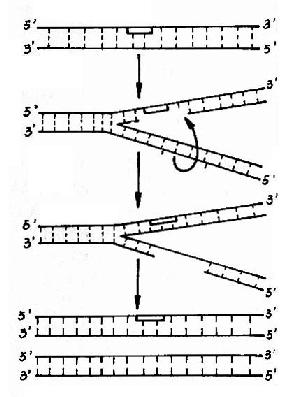
图16-22 DNA损伤后重组修复
(三)重组修复(recombinational repair)
上述的切除修复在切除损伤段落后是以原来正确的互补链为模板来合成新的段落而做到修复的。但在某些情况下没有互补链可以直接利用,例如在DNA复制进行时发生DNA损伤,此时DNA两条链已经分开,其修复可用图16?2所示的DNA重组方式:①受损伤的DNA链复制时,产生的子代DNA在损伤的对应部位出现缺口。②另一条母链DNA与有缺口的子链DNA进行重组交换,将母链DNA上相应的片段填补子链缺口处,而母链DNA出现缺口。③以另一条子链DNA为模板,经DNA聚合酶催化合成一新DNA片段填补母链DNA的缺口,最后由DNA连接酶连接,完成修补。
重组修复不能完全去除损伤,损伤的DNA段落仍然保留在亲代DNA链上,只是重组修复后合成的DNA分子是不带有损伤的,但经多次复制后,损伤就被“冲淡”了,在子代细胞中只有一个细胞是带有损伤DNA的。
(四)SOS修复
“SOS”是国际上通用的紧急呼救信号。SOS修复是指DNA受到严重损伤、细胞处于危急状态时所诱导的一种DNA修复方式,修复结果只是能维持基因组的完整性,提高细胞的生成率,但留下的错误较多,故又称为错误倾向修复(error?prone repair),使细胞有较高的突变率。
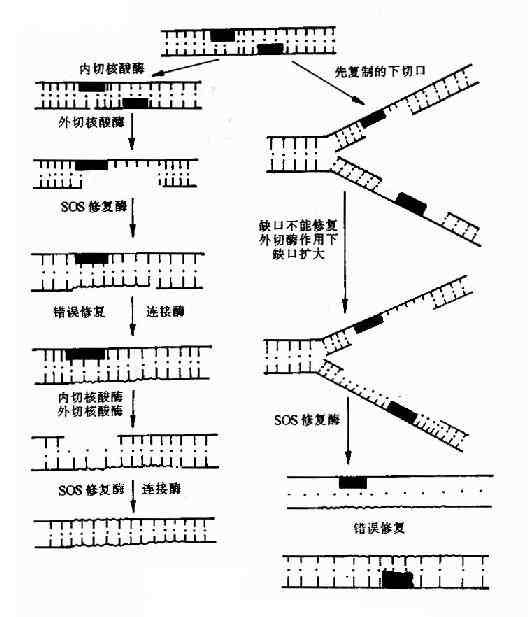
图16-23 SOS修复
如图16-23所示,当DNA两条链的损伤邻近时,损伤不能被切除修复或重组修复,这时在核酸内切酶、外切酶的作用下造成损伤处的DNA链空缺,再由损伤诱导产生的一整套的特殊DNA聚合酶桽OS修复酶类,催化空缺部位DNA的合成,这时补上去的核苷酸几乎是随机的,但仍然保持了DNA双链的完整性,使细胞得以生存。但这种修复带给细胞很高的突变率。
应该说目前对真核细胞的DNA修复的反应类型、参与修复的酶类和修复机制了解还不多,但DNA损伤修复与突变、寿命、衰老、肿瘤发生、辐射效应、某些毒物的作用都有密切的关系。人类遗传性疾病已发现4000多种,其中不少与DNA修复缺陷有关,这些DNA修复缺陷的细胞表现出对辐射和致癌剂的敏感性增加。例如着色性干皮病(xerodermapigmentosum)就是第一个发现的DNA修复缺陷性遗传病,患者皮肤和眼睛对太阳光特别是紫外线十分敏感,身体暴光部位的皮肤干燥脱屑、色素沉着、容易发生溃疡、皮肤癌发病率高,常伴有神经系统障碍,智力低下等,病人的细胞对嘧啶二聚体和烷基化的清除能力降低。
小结
环境和生物体内的因素都经常会使DNA的结构发生改变。DNA的复制会发生碱基的配对错误;体内DNA会有自发性结构变化,包括DNA链上的碱基异构互变、脱氨基、碱基修饰、DNA链上的碱基脱落等。外界射线的照射等物理因素,烷化剂、碱基类似物、修饰剂等化学因素都能损伤DNA的结构,变化包括有相邻嘧啶共价二聚体的形成、碱基、脱氧核糖和磷酸基团的烷基化和其它修饰、碱基脱落、DNA单链断裂、双链断裂、DNA链内交联、链间交联、DNA与周围的蛋白质交连等。最后能导致DNA的点突变、DNA核苷酸的缺失、插入或转位、DNA链的断裂等,结果可能影响生物细胞的功能和遗传特性,这些改变可能会导致细胞死亡、也有机会使细胞获得新的功能或进化,也可能细胞只有DNA结构的遗传性改变而没有表型变化,视DNA结构变化的部位、类型和范围不同而异。
生物在进化过程中获得的DNA修复功能,对生物的生存和维持遗传的稳定性是至关重要的。对有些DNA的损伤,细胞能将其完全修复到原样,如可将嘧啶二聚体切开、DNA单链断裂可重新连接、碱基缺失可再配对插入、加成的烷基可以移除、一条链上的碱基或核苷酸的错误可以切除并依赖互补链作模板而复制重新修复等。对DNA较严重的损伤,细胞可采取重组修复、SOS修复等方式进行反应,以期提高细胞的存活率,但不能完全消除DNA的损伤,会带给细胞较高的突变率。
DNA的损伤和修复与遗传、突变、寿命、衰老、辐射效应、肿瘤发生、某些毒剂的作用、以及某些遗传性疾病等有密切的关系。目前对DNA损伤修复的认识还不透彻。
复习思考题
1.为什么说细胞对DNA损伤的修复能力对细胞的生存是至关重要的?
2.体内那些因素会导致DNA结构的变化?细胞能采取那些办法保持DNA结构的稳定性?
3.那些环境因素容易损伤生物体内的DNA?损伤有那些方式和类型?结果对生物细胞会有些什么影响?
4.现在所知生物细胞对DNA损伤修复有那些方式?修复反应的结果会如何?

 中医世家
中医世家 浦 标 网
浦 标 网 河南大学精品课程
河南大学精品课程 图书资料室
图书资料室