病毒(Virus)是一类非细胞形态的微生物。主要有以下列基本特征:(1)个体微小,可通过除菌滤器,大多数病毒必须用电镜才能看见;(2)仅具有一种类型的核酸,或DNA或RNA;(3)严格的活细胞(真核或原核细胞)内复制增殖;(4)具有受体连结蛋白(receptor binding protein),与敏感细胞表面的病毒受体连结,进而感染细胞。病毒与其他微生物的主要区别见表21-1。
表21-1 病毒与其他微生物的主要区别要点
在无活细胞的培养基中生长
| 微生物种类 |
二等分裂繁殖 |
核酸类型* |
核糖体 |
敏感性 |
|
| 抗生素 |
干扰素 |
|
| 细菌 支原体 立克次体 衣原体 病毒 |
+ + - - - |
+ + + + -(复制) |
D+R D+R D+R D+R D或R |
+ + + + - |
+ + + + - |
- - - + + |
*D=DNA(脱氧核糖核酸);R=RNA(核糖核酸)
**有些细菌与立克次氏体对干扰素也敏感。
第一节 病毒的形态与结构
一、病毒的大小与形态
病毒个体微小,测量病毒大小的单位是毫微米(nm),即1/1000微米。在型病毒(如牛痘苗病毒)约200~300nm;中型病毒(如流感病毒)约100nm;小型病毒(如脊髓灰质炎病毒)仅20~30nm。研究病毒大小可用高分辩率电子显微镜,放大几万到几十万倍直接测量;也可用分级过滤法,根据它可通过的超滤膜孔径估计其大小;或用超速离心法,根据病毒大小,形状与沉降速度之间的关系,推算其大小。
一个成熟有感染性的病毒颗粒称“病毒体”(Viron)。电镜观察有五种形态(图21-1);
1.球形 (Sphericity)大多数人类和动物病毒为球形,如脊髓灰质炎病毒、疱疹病毒及腺病毒等。
2.丝形 (Filament)多见于植物病毒,如烟草花叶病病毒等。人类某些病毒(如流感病毒)有时也可形成丝形。
3.弹形(Bullet-shape)形似子弹头,如狂犬病病毒等,其他多为植物病毒。
4.砖形(Brick-shape)如痘病毒(无花病毒、牛痘苗病毒等)。其实大多数呈卵圆形或“菠萝形”。
5.蝌蚪形(Tadpole-shape)由一卵圆形的头及一条细长的尾组成,如噬菌体。
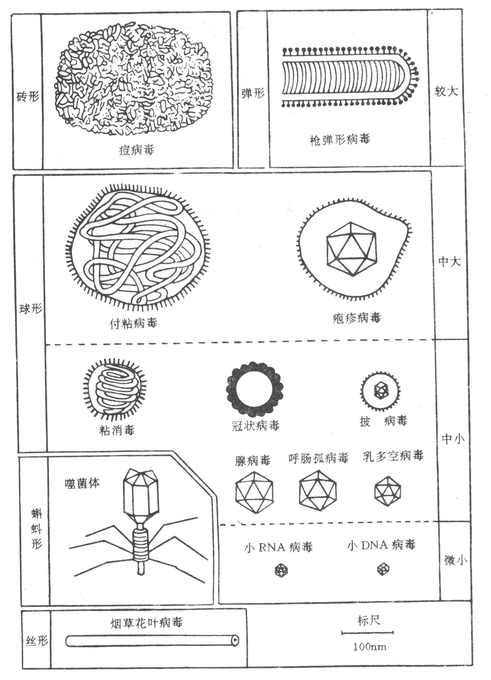
图21-1 各种主要病毒的形态与大小比较(模式图)
二、病毒的结构与功能
病毒的结构有二种,一是基本结构,为所有病毒所必备;一是辅助结构,为某些病毒所特有。它们各有特殊的生物学功能。
(一)病毒的基本结构
1.含RNA的称为RNA病毒。DNA病毒核酸多为双股(除微小病毒外),RNA病毒核酶酸多为单股(除呼肠孤病毒外)。
病毒核酸也称基因组(Genome),最大的痘病毒(Poxvirus)含有数百个基因,最小的微小病毒(Parvovirus)仅有3-4个基因。根据核酸构形及极性可分为环状、线状、分节段以及正链、负链等不同类型,对进一步阐明病毒的复制机理和病毒分类有重要意义。
核酸蕴藏着病毒遗传信息,若用酚或其他蛋白酶降解剂去除病毒的蛋白质衣壳,提取核酸并转染或导入宿主细胞,可产生与亲代病毒生物学性质一致的子代病毒,从而证实核酸的功能是遗传信息的储藏所,主导病毒的生命活动,形态发生,遗传变异和感染性。
2.衣壳(Capsid )在核酸的外面紧密包绕着一层蛋白质外衣,即病毒的“衣壳”。衣壳是由许多“壳微粒(Capsomere)”按一定几何构型集结而成,壳微米在电镜下可见,是病毒衣壳的形态学亚单位,它由一至数条结构多肽能成。根据壳微粒的排列方式将病毒构形区分为:①立体对称(Cubic symmetry),形成20个等边三角形的面,12个顶和30条棱,具有五、三、二重轴旋转对称性(图21-2),如腺病毒、脊髓灰质炎病毒等;②螺旋对称(Helical symmetry),壳微粒沿螺旋形盘红色的核酸呈规则地重复排列,通过中心轴旋转对称,图21-3,如正粘病毒,副粘病毒及弹状病毒等;③ 复合对称 (Complex symmetry),同时具有或不具有两种对称性的病毒,如痘病毒与噬菌体(图21-1)。

图21-2 腺病毒结构的模式图
蛋白质衣壳的功能是:(1)致密稳定的衣壳结构除赋予病毒固有的形状外,还可保护内部核酸免遭外环境(如血流)中核酸酶的破坏;(2)衣壳蛋白质是病毒基因产物,具有病毒特异的抗原性,可刺激机体产生抗原病毒免疫应答;(3)具有辅助感染作用,病毒表面特异性受体边连结蛋白与细胞表面相应受体有特殊的亲和力,是病毒选择性吸附宿主细胞并建立感染灶的首要步骤。
病毒的核酸与衣壳组成核衣壳(Nucleocapsid),最简单的病毒就是裸露的核衣壳,如脊髓灰质炎病毒等。有囊膜的病毒核衣壳又称为核心(core)。
(二)病毒的辅助结构
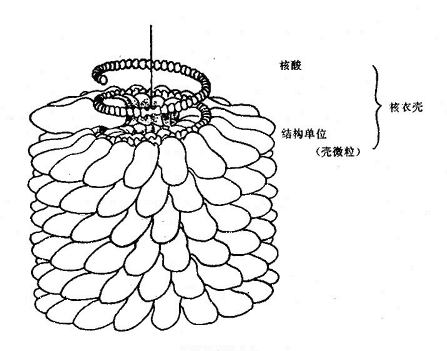
图21-3 螺旋对称病毒颗粒的核衣壳
1.囊膜(Envelope)某些病毒,如虫媒病毒、人类免疫缺陷病毒、疱疹病毒等,在核衣壳外包绕着一层含脂蛋白的外膜,称为“囊膜”。囊膜中含有双层脂质、多糖和蛋白质,其中蛋白质具有病毒特异性,常与多糖构成糖蛋白糖蛋白亚单位,嵌合在脂质层,表面呈棘状突起,称“剌突(Spike)或囊微粒(Peplomer)”。它们位于病毒体的表面,有高度的抗原性,并能选择性地与宿主细胞受体结合,促使病毒囊膜与宿主细胞膜融合,感染性核衣壳进入胞内而导致感染。囊膜中的脂质与宿主细胞膜或核膜成分相似,证明病毒是以“出芽”方式,从宿主细胞内释放过程中获得了细胞膜或核膜成分。有囊膜病毒对脂溶剂和其他有机溶剂敏感,失去囊膜后便丧失了感染性。
2.触须样纤维(Fiber)腺病毒是唯一具有触须样纤维的病毒,腺病毒的触须样纤维是由线状聚合多肽和一球形末端蛋白所组成,位于衣壳的各个顶角(图21-2)。该纤维吸附到敏感细胞上,抑制宿主细胞蛋白质代谢,与致病作用有关。此外,还可凝集某些动物红细胞。
3.病毒携带的酶某些病毒核心中带有催化病毒核酸合成的酶,如流感病毒带有RNA的RNA聚合酶,这些病毒在宿主细胞内要靠它们携带的酶合成感染性核酸。
了解病毒的形态结构、化学组成及功能,不仅对病毒的分类和鉴定有重要意义,同时也有助于理解病毒的宿主范围,致病作用及亚单位疫苗的研制。
第二节 病毒的增殖
病毒体在细胞外是处于静止状态,基本上与无生命的物质相似,当病毒进入活细胞后便发挥其生物活性。由于病毒缺少完整的酶系统,不具有合成自身成份的原料和能量,也没有核糖体,因此决定了它的专性寄生性,必须侵入易感的宿主细胞,依靠宿主细胞的酶系统、原料和能量复制病毒的核酸,借助宿主细胞的核糖体翻译病毒的蛋白质。病毒这种增殖的方式叫做“复制(Replication)”。病毒复制的过程分为吸附、穿入、脱壳、生物合成及装配释放五个步骤,又称复制周期(Replication cycle)。
一、吸附
吸附(Adsorption)是指病毒附着于敏感细胞的表面,它是感染的起始期。特异性吸附是非常重要的,根据这一点可确定许多病毒的宿主范围,不吸附就不能引起感染。细胞与病毒相互作用最初是偶然碰撞和静电作用,这是可逆的联结。脊髓灰质炎病毒的细胞表面受体是免疫球蛋白超家族,在非灵长类细胞上没有发现此受体,而猴肾细胞、Hela细胞和人二倍体纤维母细胞上有它的受体,故脊髓来质炎病毒能感染人体鼻、咽、肠和脊髓前角细胞,引起脊髓灰质炎(小儿麻痹)。水磨石病毒的细胞表面受体是含唾液酸(N-乙酰神经氨酸)的糖蛋白,它与流感病毒表面的血凝素剌突(受体连结蛋白)有特殊的亲和力,如用神经氨酸酶破坏该受体,则流感病毒不再吸附这种细胞。此外,HIV受体为CD4;鼻病毒的受体为细胞粘附分子-1(1CAM-1);EB病毒的受体为补体受体-2(CR-2)。病毒吸附也受离子强度、pH、温度等环境条件的影响。研究病毒的吸附过程对了解受体组成、功能、致病机理以及探讨抗病毒治疗有重要意义。
二、穿入
穿入(Penetration)是指病毒核酸或感染性核衣壳穿过细胞进入胞浆,开始病毒感染的细胞内期。主要有三种方式:(1)融合(Fusion),在细胞膜表面病毒囊膜与细胞膜融合,病毒的核衣壳进入胞浆。副粘病毒以融合方式进入,如麻疹病毒、腮腺炎病毒囊膜上有融合蛋白,带有一段疏水氨基酸,介导细胞膜与病毒囊膜的融合。(2)胞饮(Viropexis),由于细胞膜内陷整个病毒被吞饮入胞内形成囊泡。胞饮是病毒穿入的常见方式,也是哺乳动物细胞本身具有一种摄取各种营养物质和激素的方式。当病毒与受体结合后,在细胞膜的特殊区域与病毒病毒一起内陷形成膜性囊泡,此时病毒在胞浆中仍被胞膜覆盖。某些囊膜病毒,如流感病毒借助病毒的血凝素(HA)完成脂膜间的融合,囊泡内低Ph环境使HA蛋白的三维结构发生变化,从而介导病毒囊膜与囊泡膜的融合,病毒核衣壳进入胞浆。(3)直接进入,某些无囊膜病毒,如脊髓灰质炎病毒与受体接角后,衣壳蛋白的多肽构形发生变化并对蛋白水解酶敏感,病毒核酸可直接穿越细胞膜到细胞浆中,而大部分蛋白衣壳仍留在胞膜外,这种进入的方式较为少见。
三、脱壳
穿入和脱壳是边续的过程,失去病毒体的完整性被称为“脱壳(Uncoating)”。人脱壳到出现新的感染病毒之间叫“隐蔽期”。经胞饮进入细胞的病毒,衣壳可被吞噬体中的溶酶体酶降解而去除。有的病毒,如脊髓灰质炎病毒,在吸附穿入细胞的过程中病毒的RNA释放到胞浆中。而痘苗病毒当其复杂的核心结构进入胞浆中后,随之病毒体多聚酶活化,合成病毒脱壳所需要的酶,完成脱壳。
四、生物合成
DNA病毒的RNA病毒在复制的生化方面有区别,但复制的结果都是合成核酸分子和蛋白质衣壳,然后装配成新的有感染性的病毒。一个复制周期大约需6~8小时。
(一)双股DNA病毒的复制
多数DNA病毒为双股DNA。
双股DNA病毒,如单纯疹病毒和腺病毒在宿主细胞核内的RNA聚合酶作用下,从病毒DNA上转录病毒mRNA,然后转移到胞浆核糖体上,指导合成蛋白质。而痘苗病毒本身含有RNA聚合酶,它可在胞浆中转录mRNA。mRNA有二种:早期m RNA,主要合成复制病毒DNA所需的酶,如依赖DNA的DNA聚合酶,脱氧胸腺嘧啶激酶等,称为早期蛋白;晚期mRNa ,在病毒DNA复制之后出现,主要指导合成病毒的结构蛋白,称为晚期蛋白。
子代病毒DNA的合成是以亲代DNA为模板,按核酸半保留形式复制子代双股DNA。DNA复制出现在结构蛋白合成之前。
(二)单股RNA病毒的复制
RNA病毒核酸多为单股,病毒全部遗传信息均含在RNA中。根据病毒核酸的极性,将RNA病毒分为二组:病毒RNA的硷基序列与mRNA完全相同者,称为正链RNA病毒。这种病毒RNA可直接起病毒mRNA的作用,附着到宿主细胞核糖体上,翻译出病毒蛋白。从正链RNA病毒颗粒中提取出RNA,并注入适宜的细胞时证明有感染性;病毒RNA硷基序列与mRNA互补者,称为负链RNA病毒。负链RNA病毒的颗粒中含有依赖RNA的RNA多聚酶,可催化合成互补链,成为病毒mRNA,翻译病毒蛋白。从负链RNA病毒颗粒中提取出的RNA,因提取过程损坏了这种酶,从而无感染性。
1.正链RNA病毒的复制以脊髓灰质炎病毒为例,侵入的RNA直接附着于宿主细胞核糖体上,翻译出大分子蛋白,并迅速被蛋白水解酶降解为结构蛋白和非结构蛋白,如依赖RNA的RNA聚合酶。在这种酶的作用下,以亲代RNA为模板形成一双链结构,称“复制型(Replicativeform)”。再从互补的负链复制出多股子代正链RNA,这种由一条完整的负链和正在生长中的多股正链组成的结构,秒“复制中间体(Replicative intermediate) ”。新的子代RNA分子在复制环中有三种功能:(1)为进一步合成复制型起模板作用;(2)继续起mRNA作用;(3)构成感染性病毒RNA。
2.负链RNA病毒的复制流感病毒、副流感病毒、狂犬病毒和腮腺炎病毒等有囊膜病毒属于这一范畴。病毒体中含有RNA的RNA聚合酶,从侵入链转录出mRNA,翻译出病毒结构蛋白和酶,同时又可做为模板,在依赖RNA的RNA聚合酶作用下合成子代负链RNA。
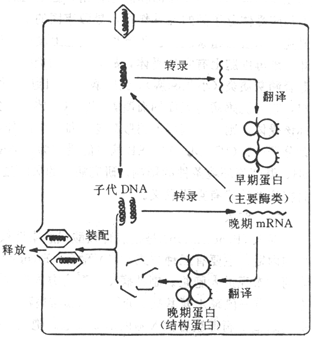
图21-4 DNA病毒复制的主要步骤
(三)逆转录病毒的复制
逆转录病毒(Retrovirus)又称RNA肿瘤病毒(Oncornavirus) ,病毒体含有单股正链RNA、依赖RNA的DNA多聚酶(逆转录酶)和tRNA。其复制过程分二个阶段:第一阶段,病毒核时进入胞浆后,以RNA为模板,在依赖RNA的DNA多聚酶和tRNA引物的作用下,合成负链DNA(即RNA:DNA),正链RNA被降解,进而以负链DNA为模板形成双股DNA(即DNA:DNA),转入细胞核内,整合成宿主DNA中,成为前病毒。第二阶段,前病毒DNA转录出病毒mRNA,翻译出病毒蛋白质。同样从前病毒DNA转录出病毒RNA,在胞浆内装配,以出芽方式释放。被感染的细胞仍持续分裂将前病毒传递至子代细胞。
(四)病毒蛋白的合成与修饰
病毒mRNA在宿主细胞聚核糖体上翻译合成病毒结构蛋白和非结构蛋白,结构蛋白是病毒结构的组成成分,非结构蛋白虽然不是病毒的结构成分,但是在病毒复制中具有重要功能,大多是一些催化、调节病毒复制的酶类和调控蛋白。
通常动物病毒mRNA仅翻译一条连续的完整的病毒多肽链,这种mRNA叫做单顺反子mRNA (Monocistronic mRNA) 。分段基因级病毒,如流感病毒,核酸分为8个节段,每一节段转录一条mRNA,翻译一种病毒蛋白。有的病毒,如脊髓灰质炎病毒,病毒RNA本身做为,mRNA,首先翻译出一大分子蛋白,然后在特殊位点被细胞或病毒蛋白水解酶裂解为许多小分子病毒蛋白,包括结构蛋白和非结构蛋白。也有的病毒,如披膜病毒,基因组上有多处转录起始和终止码,分别转录出单顺反子mRNA并合成各自的病毒蛋白。DNA的转录发生在细胞核内,转录产物经剪切拼接,并在3'端聚腺苷酸化,5'端加上甲基化帽,转送入胞浆,合成病毒蛋白。
某些病毒蛋白合成后需要修饰,如磷酸化、糖基化等。由病毒和细胞的蛋白激酶完成磷酸化,这是活化或灭活某些蛋白的一种方式。病毒糖蛋白是在胞浆中与膜相连的核糖体上合成,经粗面内质网、平滑内质网、高尔基氏体到达细胞膜,在此过程中被糖基化。
五、装配与释放
新合成的病毒核酸和病毒结构蛋白在感染细胞内组合成病毒颗粒的过程称为装配(Assembly),而从细胞内转移到细胞外的过程为释放(Release)。大多数DNA病毒,在核内复制DNA,在胞浆内合成蛋白质,转入核内装配成熟。而痘苗病毒其全部成份及装配均在胞浆内完成。RNA病毒多在胞浆内复制核酸及合成蛋白。感染后6个小时,一个细胞可产生多达10,000个病毒颗粒。
病毒装配成熟后释放的方式有:(1)宿主细胞裂解,病毒释放到周围环境中,见于无囊膜病毒,如腺病毒、脊髓灰质炎病毒等;(2)以出芽的方式释放,见于有囊膜病毒,如疱疹病毒在核膜上获得囊膜,流感病毒在细胞膜上获得囊膜而成熟,然后以出芽方式释放出成熟病毒。也可通过细胞间桥或细胞融合邻近的细胞。
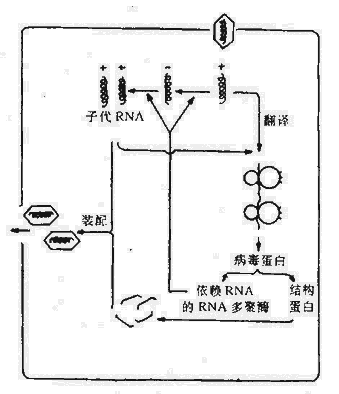
图21-5 正链RNA病毒复制的主要步骤
病毒的增殖不只是产生有感染性的子代,绝大多数动物病毒在大量感染的情况下,经多次增殖会产生缺损干扰颗粒(Defective interfering particles),它是能干扰亲代病毒复制的缺损病毒,其核酸有部分缺损或被宿主DNA片段替换。缺损干扰颗粒的基本特性是:(1)本身不能繁殖;(2)有辅助病毒存在时方能增殖;(3)干扰同种病毒而不干扰异种病毒的增殖;(4)在感染细胞内与亲代病毒竞争性增殖。由于缺损干扰颗粒的产生,使同种感染性病毒数量减少,在导致病毒的持续性感染中具有一定的作用,但疫苗中含有大量缺损干扰颗粒会影响活疫苗的免疫效果。
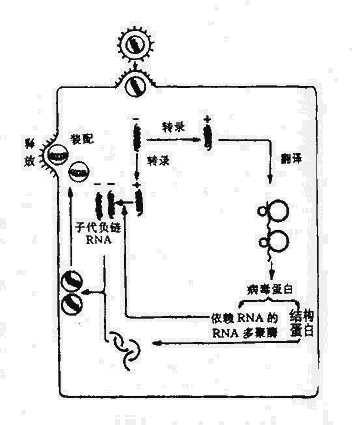
图21-6 负链RNA病毒复制的主要步骤
第三节 抵抗力与变异
一、病毒对理化因素的抵抗力
(一)物理因素
1.温度大多数病毒(除肝炎病毒外)耐冷而不耐热。病毒一旦离开机体,经加热56~60℃30分钟,由于表面蛋白变性,而丧失其感染性,即被灭活。病毒对低温的抵抗力较强,通常在-20~196℃仍不失去活性,但对反复冻融则敏感。一般可用低温真空干燥法(Lyophilization)保存病毒,但在室温条件下干燥易使病毒灭活。
2.盐类对病毒的稳定作用克分子浓度的盐可提高病毒对热的抵抗力。MgCl2对脊液灰质炎病毒、MgSO4对正粘和副粘病毒、Na2SO4对疱疹病毒具有稳定作用。因此在减毒活疫苗中须加这类稳定剂。有囊膜病毒即使在-90℃也不能长期保存,但加入保护剂如二甲基亚砜(DMSO)可使之稳定。
3. pH 病毒一般在pH5.0~9.0的环境是稳定的,但在某些病毒的血凝反应中,pH改变可影响改变试验的结果。
4.射线 紫外线、X线和高能量粒子可杀活病毒,这是因为光量子可击毁病毒核酸的分子结构,不同病毒其敏感度不一。
某些活性染料(如甲苯胺兰、中性红、丫啶橙)对病毒具有不同程度的渗透作用,这些染料与病毒核酸结合后,易被可见光灭活。
(二)化学因素
1.脂溶剂有囊膜病毒可迅速被脂溶剂破坏,如乙醚、氯彷、去氧胆酸钠。这类病毒通常不能在含有胆汁的肠道中引起感染。病毒对脂溶剂的敏感性可作为病毒分类的依据之一。
2.甘油 大多数病毒在50%甘油盐水中能活存较久。因病毒体中含游离水,不受甘油脱水作用的影响,故可用于保存病毒感染的组织。
3.化学消毒剂一般病毒对高锰酸钾、次氯酸盐等氧化剂都很敏感,升汞、酒精、强酸及强碱均能迅速杀灭病毒,但0.5%~1%石炭酸仅对少数病毒有效。饮水中漂白粉浓度对乙型肺炎,肠道病毒无效。β-丙内脂(β-Propiolactone)及环氧乙烷(Ethylene oxide)可杀灭各种病毒。
4.抗生素抗生素及磺胺对病毒无效。利福平(Rifampin)能抑制痘病毒复制,干扰病毒DNA或RNA合成,但也干扰宿主细胞的代谢,有较强的细胞毒性作用。
二、病毒的变异
(一)突变
病毒的突变(Mutation)是指基因组中核酸硷基顺序上的化学变化,可以是一个核苷酸的改变,也可为上百上千个核苷酸的缺失或易位。病毒复制中的自然突变率10-5~10-8,而各种物理、化学诱变剂(Mutagens)可提高突变率,如温度、射线、5-溴尿嘧啶、亚硝酸盐等的作用均可诱发突变。突变株与原先的野生型病毒(Wild-type virus)特性不同,表现为病毒毒力、抗原组成、温度和宿主范围等方面的改变。
1.毒力改变有强毒株及弱毒株,后者可制成弱毒活病毒疫苗,如脊液灰质炎疫苗、麻疹疫苗等。
2.条件致死突变株指病毒突变后在特定条件下能生长,而在原来条件下不能繁殖而被致死。其中最主要是的是温度敏感条件致死突变株(Temperature-sensitive conditional lethalmutant),简称温度敏感突变株(ts株),在特定温(28~35℃)下孵育则能增殖,在非特定温度(37~40℃)下孵育则不能繁殖,而野生型在两种温度均能增殖。显然是由于在非特定温度下,突变基因所编码的蛋白缺乏其应有功能。因此大多数ts株同时又是减毒株。现已从许多动物病毒中分离出ts株,选择遗传稳定性良好的品系用于制备碱毒活疫苗,如流感病毒及脊髓灰制裁炎病毒ts 株疫苗。
3.宿主适应性突株例如狂犬病毒突变株适应在兔脑内增殖,由“街毒”变为“固定毒”,可制成狂犬病疫苗。
(二)基因重组
当二种有亲缘关系的不同病毒感染同一宿主细胞时,它们的遗传物质发生交换,结果产生不同于亲代的可遗传的子代,称为基因重组(Genetic recombination)。
1.活病毒间的重组 例如流感病毒两个亚型之间可基因重组,产生新的杂交株,即具有一个亲代的血凝素和另一亲代的神经氨酸酶。这在探索自然病毒变异原理中具有重要意义。流感每隔十年左右引起一次世界性大流行,可能是由于人的流感病毒与某些动物(鸡、马、猪)的流感病毒间发生基因重组所致。
2.灭活病毒间的重组例如用紫外线灭活的两株同种病毒,若一同培养后,常可使灭活的病毒复活,产生出感染性病毒体,此称为多重复活(Multiplicity reactivation),这是因为两种病毒核酸上受损害的基因部位不同,由于重组合相互弥补而得到复活。因此现今不用紫外线灭活病毒制造疫苗,以防病毒复活的危险。
3.死活病毒间的重组例如将能在鸡胚中生长良好的甲型流感病毒(A0或A1亚型)疫苗株经紫外线灭活后,再加亚洲甲型(A2亚型)活流感病毒一同培养,产生出具有前者特点的A2亚型流感病毒,可供制作疫苗,此称为交叉复活(Cross reactivation)。
(三)基因产物的相互作用
1.表型混合(Phenotypemixing)两种病毒混合感染后,一个病毒的基因组偶而装入另一病毒的衣壳内,或装入两个病毒成分构成的衣壳内,发生表型混合。这种混合是不稳定的,传代后可恢复其原来的特性。
2.基因型混合(Genotypemixing)指两种病毒的核酸偶而混合装在同一病毒衣壳内,或两种病毒的核衣壳偶尔包在一个囊膜内,但它们的核酸都未重组合,所以没有遗传性。
3.互补(Complementation)指两种病毒通过其产生的蛋白质产物(如酶、衣壳或囊膜)相互间补助不足,例如辅助病毒与缺损病毒间、两个缺损病毒间、活病毒与死病毒间都可以互补,互补后仍产生原来病毒的子代。
4.增强(Enhancement)指两种病毒混合培养时,一种病毒能促进增强另一种病毒的产量,可能是因为前者压制了产生干扰素所致。
(四)病毒变异的实际意义
1.研制减毒活疫苗 如ts株、宿主适应性突变株的研制。
2.应用于基因工程(Geneticengineering)基因工程是将一个生物体的基因(Gene),也就是携带遗传信息的DNA片段,转移到另一个生物内,与原有生物体的DNA结合,实现遗传性状的转移和重新组合,从而使人们能够定向地控制、干予和改变生物体的变异和遗传。目前病毒基因工程正沿着二个方向发展:一是将编码病毒表面抗原的基因移植到质粒中去,在大肠杆菌中产生大量表面抗原物质,以制备疫苗或诊断用抗原。如乙型肺炎病毒编码表面抗原的DNA片段已在酵母菌中表达,该疫苗正进行人体观察;二是探索病毒作为基因工程载体的可能性,以便将所需要的外源基因带入人体或支物体内,以治疗人类遗传疾病或创造动物新品种的目的。
[附]病毒的分类与命名
按生物分类学标准,分类应能反映生物体的进化与种系发生的关系。但病毒不可能按这个原则分类,一般仍按病毒鉴定等的实际需要而分类。
各种生物体均可有病毒寄生。已知有动物病毒、植物病毒、放线菌病毒及细菌病毒(即噬菌体)。医学上重要的病毒有500种左右,其中有不少对人类有致病力。有些病毒也能在节肢动物中增殖(如虫媒病毒)。目前通过的分类法是将支物病毒分为脱氧核糖病毒(Deoxyriboviruses 简称DNA病毒)与核糖核酸病毒(Riboviruses 简称RNA病毒 )两大类。再按病毒的理化性状分为若干科(族属)(表21-2)。
表21-2 按病毒理化及生物性状的分类(供参考)
| 核心中的核酸 |
衣壳的对称性 |
囊膜 |
对乙醚敏感性 |
壳微粒数 目 |
病毒体大小(nm③) |
核酸分子量(×106) |
核酸物理学形式 |
基因数(约数) |
病毒科名 |
| DNA病毒 |
20面体 对 称 |
- |
不敏感 |
32 72 252 |
18~26 45~55 70~90 |
1.5~1.8 3~5 20~30 |
单 股 双股环状 双 股 |
3~4 5~8 30 |
小DNA病毒科(Parvoviridee) 乳头多瘤空泡病毒科(Papovaviridae) 腺病毒科(Adenoviridae) |
| + |
敏感 |
162 ? |
100④ 42 |
90~130 2.1 |
双 股 环状双股 |
160 7 |
疱疹病毒科(Herpetoviridae) 嗜肝病毒科(Hepadnaviridae) |
| 复合 对称 |
复合外壳 |
① |
|
230~300 |
130~240 |
双 股 |
300 |
痘病毒科(Poxviridae) |
| RNA病毒 |
20面体对称 |
- |
不敏感 |
32 ?② |
20~30 60~80 |
2~2.8 12~19 |
单 股 双股分段 |
4~6 10~12 |
小核糖核酸病毒科(Picornaviridae) 吸肠弧病毒科(Reoviridae) |
| + |
敏感 |
32 |
40~70 40~50 |
3~4 3~4 |
单 股 单 股 |
10 10 |
披膜病毒科(Togaviridae) 黄病毒科(Flaviviridae) |
| 螺旋对称 |
+ |
敏感 |
|
90~100 80~120 150~300 70~175 |
6~15 5 5~8 3~4 |
单股分段 单股分段 单 股 单 股 |
23 10 >10 5 |
布尼安病毒科(Bunyaviridae) 正粘病毒科(Orthomyxoviridae) 副粘病毒科(Paramyxoviridae) 弹状病毒科(Rhabdoviridae) |
| 不明或复合对称 |
+ |
敏感 |
|
50~300 80~130 ~100 |
3~5 9 7~10 |
单股分段 单 股 单股双倍 |
10 30 20~30 |
沙粒状病毒科(Arenaviridae) 冠状病毒科(Coronaviridae) 逆转录病毒科(Retroviridae) |
说明:①正痘病毒(Orthopoxvirus)包括天花、类天花、牛痘、牛痘苗、兔痘、猴痘及鼠脱脚病等病毒皆对乙醚有抵抗性,而其他的痘病毒则对乙醚敏感。
②呼肠病毒(Reovirus)有一个外衣壳和一个内衣壳。内壳有32个壳微粒,外衣壳的壳微粒数目则未确定,认为总共有92个壳微粒。
③病毒体大小是指直径或直径×长度。
④脱去囊膜的裸露病毒体,其核衣壳的直径为100nm,但有囊膜的病毒体则可大至200nm。

 中医世家
中医世家 浦 标 网
浦 标 网 河南大学精品课程
河南大学精品课程 图书资料室
图书资料室