氨基酸是构成蛋白质分子的基本单位。蛋白质是生命活动的基础。体内的大多数蛋白质均不断地进行分解与合成代谢,细胞中不停地利用氨基酸合成蛋白质和分解蛋白质成为氨基酸。体内的这种转换过程一方面可清除异常蛋白质,这些异常蛋白质的积聚会损伤细胞。另一方面使酶或调节蛋白的活性由合成和分解得到调节,进而调节细胞代谢。实际上酶的水平取绝于其合成同样也由酶的分解来决定。所以,对细胞来说,蛋白质的分解与合成同样重要。
蛋白质分解代谢首先在酶的催化下水解为氨基酸,而后各氨基酸进行分解代谢,或转变为其它物质、或参与新的蛋白质的合成。因此氨基酸代谢是蛋白质分解代谢的中心内容。
第一节 氨基酸的一般代谢
食物蛋白经过消化吸收后,以氨基酸的形式通过血液循环运到全身的各组织。这种来源的氨基酸称为外源性基酸。机体各组织的蛋白质在组织酶的作用下,也不断地分解成为氨基酸;机体还能合成部分氨基酸(非必需氨基酸);这两种来源的氨基酸称为内源性氨基酸。外源性氨基酸和内源性氨基酸彼此之间没有区别,共同构成了机体的氨基酸代谢库(metabolic pool)。氨基酸代谢库通常以游离氨基酸总量计算,机体没有专一的组织器官储存氨基酸,氨基酸代谢库实际上包括细胞内液、细胞间液和血液中的氨基酸。
氨基酸的主要功能是合成蛋白质,也合成多肽及其他含氮的生理活性物质。除了维生素之外(维生素PP是个例外)体内的各种含氮物质几种都可由氨基酸转变而成,包括蛋白质、肽类激素、氨基酸衍生物、黑色素、嘌呤碱、嘧啶碱、肌酸、胺类、辅酶或辅基等。
从氨基酸的结构上看,除了侧链R基团不同外,均有α-氨基和α?羧基。氨基酸在体内的分解代谢实际上就是氨基、羧基和R?基团的代谢。氨基酸分解代谢的主要途径是脱氨基生成氨ammonia)和相应的α?酮酸;氨基酸的另一条分解途径是脱羧基生成CO2和胺。胺在体内可经胺氧化酶作用,进一步分解生成氨和相应的醛和酸。氨对人体来说是有毒的物质,氨在体内主要合成尿素排出体外,还可以合成其它含氮物质(包括非必需氨基酸、谷氨酰胺等),少量的氨可直接经尿排出。R?基团部分生成的酮酸可进一步氧化分解生成CO2和水,并提供能量,也可经一定的代谢反应转变生成糖或脂在体内贮存。由于不同的氨基酸结构不同,因此它们的代谢也有各自的特点。
各组织器官在氨基酸代谢上的作用有所不同,其中以肝脏最为重要。肝脏蛋白质的更新速度比较快,氨基酸代谢活跃,大部分氨基酸在肝脏进行分解代谢,同时氨的解毒过程主要也在肝脏进行。分枝氨基酸的分解代谢则主要在肌肉组织中进行。
食物中蛋白质的含量也影响氨基酸的代谢速率。高蛋白饮食可诱导合成与氨基酸代谢有关的酶系,从而使代谢加快(图7-1)。
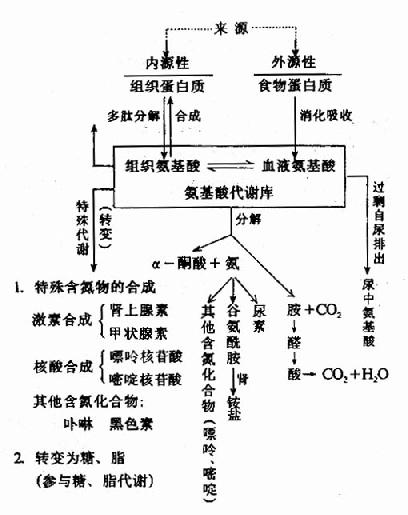
图7-1 氨基酸代谢的基本概况
一、氨基酸的脱氨基作用
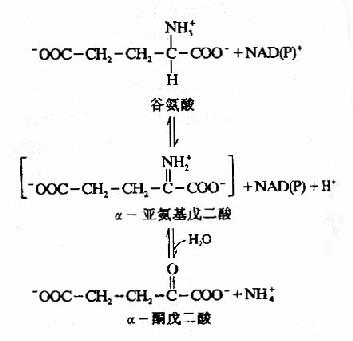
图7-2 谷氨酸脱氢酶催化的氧化脱氢反应
脱氨基作用是指氨基酸在酶的催化下脱去氨基生成α?酮酸的过程。这是氨基酸在体内分解的主要方式。参与人体蛋白质合成的氨基酸共有20种,它们的结构不同,脱氨基的方式也不同,主要有氧化脱氨、转氨、联合脱氨和非氧化脱氨等,以联合脱氨基最为重要。
(一)氧化脱氨基作用(Oxidative Deamination)
氧化脱氨基作用是指在酶的催化下氨基酸在氧化脱氢的同时脱去氨基的过程。
不需氧脱氢酶催化的氧化脱氨基作用
谷氨酸在线粒体中由谷氨酸脱氢酶(glutamatedehydrogonase)催化氧化脱氨。谷氨酸脱氢酶系不需氧脱氢酶,以NAD+或NADP+作为辅酶。氧化反应通过谷氨酸Cα脱氢转给NAD(P)+形成α?亚氨基戊二酸,再水解生成α?酮戊二酸和氨(图7-2)。
谷氨酸脱氢酶为变构酶。GDP和ADP为变构激活剂,ATP和GTP为变构抑制剂。
在体内,谷氨酸脱氢酶催化可逆反应。一般情况下偏向于谷氨酸的合成(△G°′≈30kJ·mal?1),因为高浓度氨对机体有害,此反应平衡点有助于保持较低的氨浓度。但当谷氨酸浓度高而NH3浓度低时,则有利于脱氨和α酮戊二酸的生成。
(二)转氨基作用
转氨基作用(Transamination)指在转氨酶催化下将α-氨基酸的氨基转给另一个α-是酮酸,生成相应的α?酮酸和一种新的α-氨基酸的过程。
体内绝大多数氨基酸通过转氨基作用脱氨。参与蛋白质合成的20种α-氨基酸中,除甘氨酸、赖氨酸、苏氨酸和脯氨酸不参加转氨基作用,其余均可由特异的转氨酶催化参加转氨基作用。转氨基作用最重要的氨基受体是α?酮戊二酸,产生谷氨酸作为新生成氨基酸:

进一步将谷氨酸中的氨基转给草酰乙酸,生成α?酮戊二酸和天冬氨酸:

或转给丙酮酸。生成α?酮戊二酸和丙氨酸,通过第二次转氨反应,再生出α?酮戊二酸。
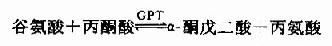
因而体内有较强的谷草转氨酸(glutamicpyruvic transaminase,GPT)和谷丙转氨酸(glutamic oxaloacetictrans aminase,GOT)活性。
转氨基作用是可逆的,该反应中△G°′≈0,所以平衡常数约为1。反应的方向取绝于四种反应物的相对浓度。因而,转氨基作用也是体内某些氨基酸(非必需氨基酸)合成的重要途径。
2.转氨基作用机理:
转氨基作用过程可分为两个阶段:
(1)一个氨基酸的氨基转到酶分子上,产生相应的酮酸和氨基化酶:
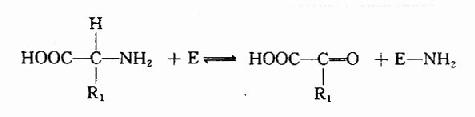
(2)NH2转给另一种酮酸,(如α?酮戊二酸)生成氨基酸,并释放出酶分子:
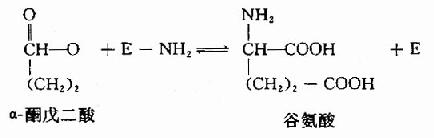
为传送NH2基因,转氨酶需其含醛基的辅酶-磷酸吡哆醛(pyridoxal-5′-phosphate,PLP)的参与。在转氨基过程中,辅酶PLP转变为磷酸吡哆胺(pyridoxamine?5′?phosphate,PMP)。PLP通过其醛基与酶分子中赖氨酸ω氨基缩合形成Schiff碱而共价结合子酶分子中。
Esmond Snell,Alexande Branstein和David Metgler等揭示转氨作用是一种兵乓机制,二阶段各分三步进行(图7-3)。
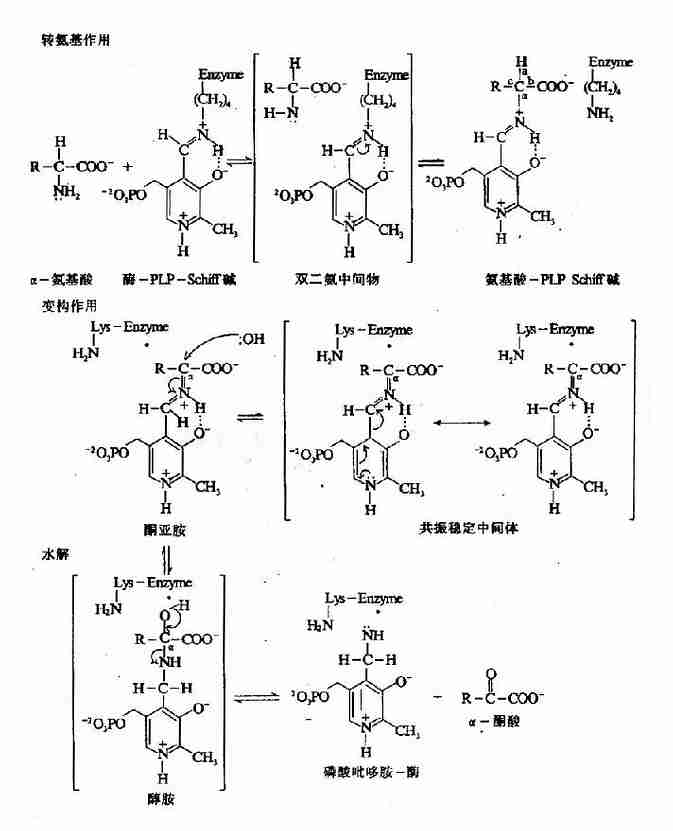
图7-3 PLP?依赖的酶促转氨基反应机理
第一阶段:氨基酸转变为酮酸
(1)氨基酸的亲核性NH2基团作用于酶?PLp Schiff碱C原子,通过转亚氨基反应(transimination ortrans?Schiffigation)形成一种氨基酸?PLp Schiff碱,同时使酶分子中赖氨酸的NH2基团复原。
(2)通过酶活性位点赖氨酸催化去除氨基酸α氢,并通过一共振稳定的中间产物在PLP第4位C原子上加质子,将氨基酸桺Lp Schiff碱分子重排为一个α?酮酸?PMP schiff碱。
(3)水解生成PMP和α-酮酸。
第二阶段:α-酮酸转变为氨基酸
为完成转氨反应循环,辅酶必需由PMP形式转变为E-PLp-Schiff形式,此过程亦包括三步,为上述反应的逆过程。
(1)PMP与一个α-酮酸作用形成α-酮酸-Schiff碱。
(2)分子重排,α-酮酸-PMp-Schiff碱变为氨基酸-PLP-Schiff碱。
(3)酶活性位点赖氨酸ω-NH2基团攻击氨基酸-PLp-Schiff碱,通过转亚氨基生成有活性的酶-PLP Schiff碱,并释放出形成的新氨基酸。
转氨基反应中,辅酶在PLP和PMP间转换,在反应中起着氨基载体的作用,氨基在α-酮酸和α-?氨基酸之间转移。可见在转氨基反应中并无净NH3的生成。
3.转氨基作用的生理意义
转氨基作用起着十分重要的作用。通过转氨作用可以调节体内非必需氨基酸的种类和数量,以满足体内蛋白质合成时对非必需氨基酸的需求。
转氨基作用还是联合脱氨基作用的重要组成部分,从而加速了体内氨的转变和运输,勾通了机体的糖代谢、脂代谢和氨基酸代谢的互相联系。
(三)联合脱氨基作用
联合脱氨基作用是体内主要的脱氨方式。主要有两种反应途径:
1.由L-谷氨酸脱氢酶和转氨酶联合催化的联合脱氨基作用:先在转氨酶催化下,将某种氨基酸的α-氨基转移到α-酮戊二酸上生成谷氨酸,然后,在L-谷氨酸脱氢酶作用下将谷氨酸氧化脱氨生成α-酮戊二酸,而α-酮戊二酸再继续参加转氨基作用。
L-谷氨酸脱氢酶主要分布于肝、肾、脑等组织中,而α-酮戊二酸参加的转氨基作用普遍存在于各组织中,所以此种联合脱氨主要在肝、肾、脑等组织中进行。联合脱氨反应是可逆的,因此也可称为联合加氨。
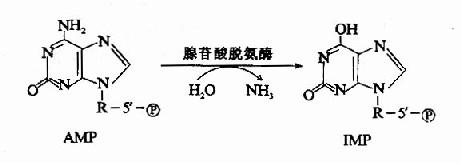
2.嘌呤核苷酸循环(purine nucleotide cycle):骨骼肌和心肌组织中L?谷氨酸脱氢酶的活性很低,因而不能通过上述形式的联合脱氨反应脱氨。但骨骼肌和心肌中含丰富的腺苷酸脱氨酶(adenylatedeaminase),能催化腺苷酸加水、脱氨生成次黄嘌呤核苷酸(IMP)。
一种氨基酸经过两次转氨作用可将α-氨基转移至草酰乙酸生成门冬氨酸。门冬氨酸又可将此氨基转移到次黄嘌呤核苷酸上生成腺嘌呤核苷酸(通过中间化合物腺苷酸代琥珀酸)。其脱氨过程可用图7-4表示。
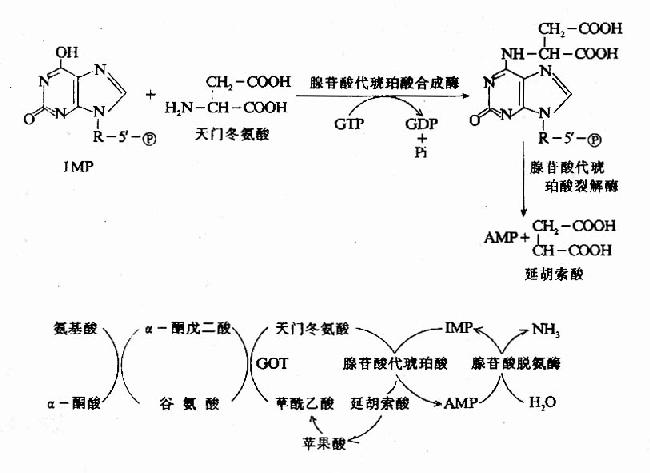
图7-4 腺嘌呤核苷酸循环
目前认为嘌呤核苷酸循环是骨骼肌和心肌中氨基酸脱氨的主要方式。John lowenstein证明此嘌呤核苷酸循环在肌肉组织代谢中具有重要作用。肌肉活动增加时需要三羧酸循环增强以供能。而此过程需三羧酸循环中间产物增加,肌肉组织中缺乏能催化这种补偿反应的酶。肌肉组织则依赖此嘌呤核苷酸循环补充中间产物-草酰乙酸。研究表明肌肉组织中催化嘌呤核苷酸循环反应的三种酶的活性均比其它组织中高几倍。AMP脱氨酶遗传缺陷患者(肌腺嘌呤脱氨酶缺乏症)易疲劳,而且运运后常出现痛性痉挛。
这种形式的联合脱氨是不可逆的,因而不能通过其逆过程合成非必需氨基酸。这一代谢途径不仅把氨基酸代谢与糖代谢、脂代谢联系起来,而且也把氨基酸代谢与核苷酸代谢联系起来。
(四)非氧化脱氨基作用(non?oxidativedeamination)
某些氨基酸还可以通过非氧化脱氨基作用将氨基脱掉。
1.脱水脱氨基 如丝氨酸可在丝氨酸脱水酶的催化下生成氨和丙酮酸。
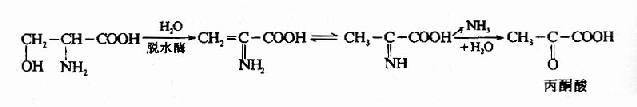
苏氨酸在苏氨酸脱水酶的作用下,生成α-酮丁酸,再经丙酰辅酶A,琥珀酰AoC参加代谢,如下图所示。
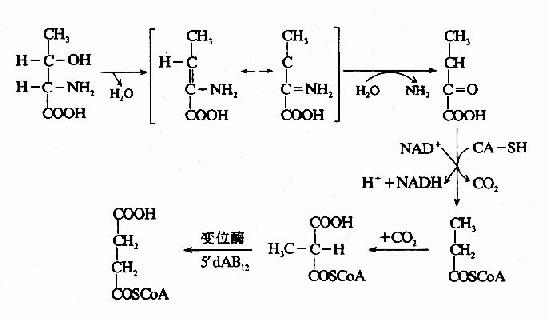
这是苏氨酸在体内分解的途径之一。
2.脱硫化氢脱氨基 半胱氨酸可在脱硫化氢酶的催化下生成丙酮酸和氨。
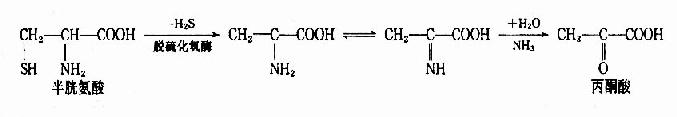
3.直接脱氨基 天冬氨酸可在天冬氨酸酶作用下直接脱氨生成延胡索酸和氨。

二、氨的代谢
(一)氨的来源
1.组织中氨基酸分解生成的氨 组织中的氨基酸经过联合脱氨作用脱氨或经其它方式脱氨,这是组织中氨的主要来源。组织中氨基酸经脱羧基反应生成胺,再经单胺氧化酶或二胺氧化酶作用生成游离氨和相应的醛,这是组织中氨的次要来源,组织中氨基酸分解生成的氨是体内氨的主要来源。膳食中蛋白质过多时,这一部分氨的生成量也增多。
2.肾脏来源的氨 血液中的谷氨酰胺流经肾脏时,可被肾小管上皮细胞中的谷氨酰胺酶(glutaminase)分解生成谷氨酸和NH3。
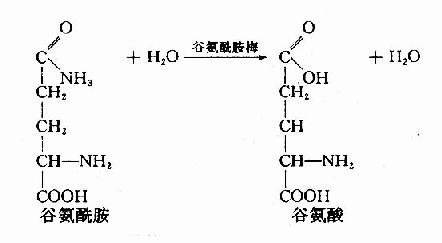
这一部分NH3约占肾脏产氨量的60%。其它各种氨基酸在肾小管上皮细胞中分解也产生氨,约占肾脏产氨量的40%。
肾小管上皮细胞中的氨有两条去路:排入原尿中,随尿液排出体外;或者被重吸收入血成为血氨。氨容易透过生物膜,而NH+4不易透过生物膜。所以肾脏产氨的去路决定于血液与原尿的相对pH值。血液的pH值是恒定的,因此实际上决定于原尿的pH值。原尿pH值偏酸时,排入原尿中的NH3与H+结合成为NH+4,随尿排出体外。若原尿的pH值较高,则NH3易被重吸收入血。临床上血氨增高的病人使用利尿剂时,应注意这一点。
3.肠道来源的氨 这是血氨的主要来源。正常情况下肝脏合成的尿素有15?0%经肠粘膜分泌入肠腔。肠道细菌有尿素酶,可将尿素水解成为CO2和NH3,这一部分氨约占肠道产氨总量的90%(成人每日约为4克)。肠道中的氨可被吸收入血,其中3/4的吸收部位在结肠,其余部分在空肠和回肠。氨入血后可经门脉入肝,重新合成尿素。这个过程称为尿素的肠肝循环(entero?hepatin circulation of urea)。
肠道中的一小部分氨来自腐败作用(putrescence)。这是指未被消化吸收的食物蛋白质或其水解产物氨基酸在肠道细菌作用下分解的过程。腐败作用的产物有胺、氨、酚、吲哚、H2S等对人体有害的物质,也能产生对人体有益的物质,如脂肪酸、维生素K、生物素等。
肠道中NH3重吸收入血的程度决定于肠道内容物的pH值,肠道内pH值低于6时,肠道内氨生成NH+4,随粪便排出体外;肠道内pH值高于6时,肠道内氨吸收入血。临床上给高血氨病人作灌肠治疗时,禁忌使用肥皂水等,以免加重病情。
(二)氨的去路
氨是有毒的物质,人体必须及时将氨转变成无毒或毒性小的物质,然后排出体外。主要去路是在肝脏合成尿素、随尿排出;一部分氨可以合成谷氨酰胺和门冬酰胺,也可合成其它非必需氨基酸;少量的氨可直接经尿排出体外。尿中排氨有利于排酸。
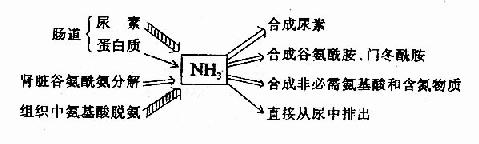
图7-5 氨的来源和去路
(三)氨的转运
1.葡萄糖-丙氨酸循环:肌肉组织中以丙酮酸作为转移的氨基受体,生成丙酸经血液运输到肝脏。在肝脏中,经转氨基作用生成丙酮酸,可经糖异生作用生成葡萄糖,葡萄糖由血液运输到肌肉组织中,分解代谢再产生丙酮酸,后者再接受氨基生成丙氨酸。这一循环途径称为“丙氨酸椘咸烟茄?贰?alanine?glucose cycle)。通过此途径,肌肉氨基酸的NH2基,运输到脏脏以NH3或天冬氨酸合成尿素。(图7-6)?
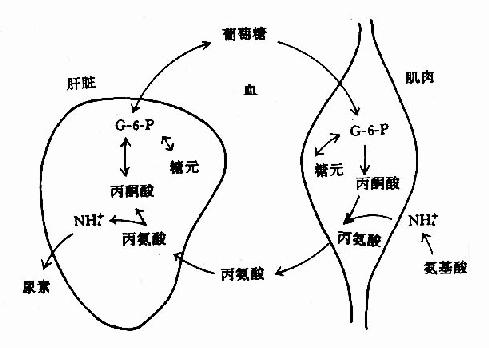
图7-6 葡萄糖?丙氨酸循环
饥饿时通过此循环将肌肉组织中氨基酸分解生成的氨及葡萄糖的不完全分解产物丙酮酸,以无毒性的丙氨酸形式转运到肝脏作为糖异生的原料。肝脏异性生成的葡萄糖可被肌肉或其它外周组织利用。
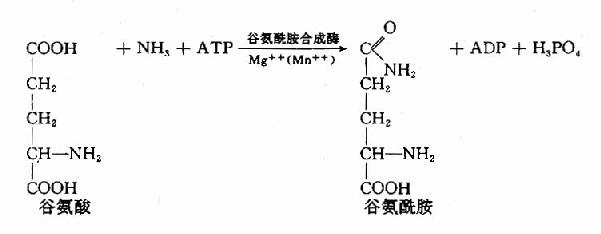
2.氨与谷氨酸在谷氨酰胺合成酶(glutamine synthetase)的催化下生成谷氨酰胺(glutamine),并由血液运输至肝或肾,再经谷氨酰酶(glutaminaes)水解成谷氨酸和氨。谷氨酰胺主要从脑、肌肉等组织向肝或肾运氨。
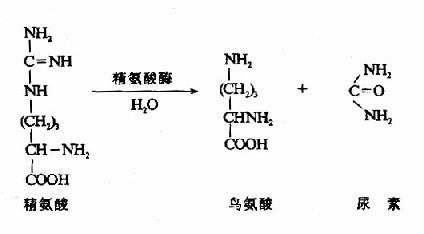
(四)尿素合成
根据动物实验,人们很早就确定了肝脏是尿素合成的主要器官,肾脏是尿素排泄的主要器官。1932年Krebs等人利用大鼠肝切片作体外实验,发现在供能的条件下,可由CO2和氨合成尿素。若在反应体系中加入少量的精氨酸、鸟氨酸或瓜氨酸可加速尿素的合成,而这种氨基酸的含量并不减少。为此,Krebs等人提出了鸟氨酸循环(ornithine cyclc)学说。其后由Ratner和Cohen详细论述了其各步反应。鸟氨酸循环可概括为:
尿素中的两个N原子分别由氨和天冬氨酸提供,而C原子来自HCO-3,五步酶促反应,二步在线粒体中,三步在胞液中进行。其详细过程可分为以下五步:
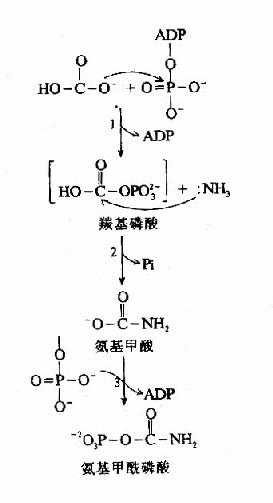
图7-7 CPS?Ⅰ 作用机理
1.氨基甲酰磷酸的合成
氨基甲酰磷酸(carbamylphosphate)是在Mg++、ATP及N?乙酰谷氨酸(N?acetyl glutamic acid,AGA)存在的情况下,由氨基甲酰磷酸合成酶I(carbamyl phosphate synthetaseI, CPS?I)催化NH3和HCO-3在肝细胞线粒体中合成。
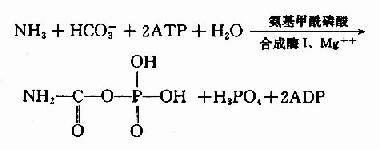
真核细胞中有两种CPS:(1)线粒体CPS?-Ⅰ利用游离NH3为氮源合成氨基甲酰磷酸,参与尿素合成。(2)胞液CPS-Ⅱ,利用谷氨酰胺作N源,参与嘧啶的从头合成。
CPS-Ⅰ催化的反应包括下述三步(图7-7)。
(1)ATP活化HCO-3生成ADP和羰基硫酸(carbonyl phosphate)
(2)NH2与羰基硫酸作用替代硫酸根,生成氨基甲酸(carbamate)和Pi。
(3)第2个ATP对氨甲酸磷酸化,生成氨基甲酰磷酸和ADP。
此反应是不可逆的,消耗2分子ATP。CPS?1是一种变构酶,AGA是此酶变构激活剂。由乙酰CoA和谷氨酸缩合而成。
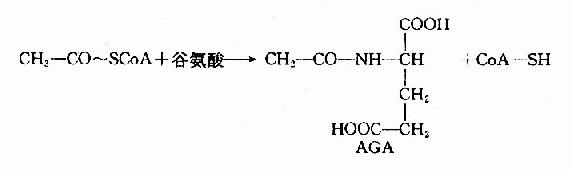
肝细胞线粒体中谷氨酸脱氢酶和氨基甲酰磷酸合成酶I催化的反应是紧密偶联的。谷氨酸脱氢酶催化谷氨酸氧化脱氨,生成的产物有NH3和NADH+H+。NADH经NADH氧化呼吸链传递氧化生成H2O,释放出来的能量用于ADP磷酸化生成ATP。因此谷氨酸脱氢酶催化反应不仅为氨基甲酰磷酸的合成提供了底物NH3,同时也提供了该反应所需要的能量ATP。氨基甲酰磷酸合成酶I将有毒的氨转变成氨基甲酰磷酸,反应中生成的ADP又是谷氨酸脱氢酶的变构激活剂,促进谷氨酸进一步氧化脱氨。这种紧密偶联有利于迅速将氨固定在肝细胞线粒体内,防止氨逸出线粒体进入细胞浆,进而透过细胞膜进入血液,引起血氨升高。
2.瓜氨酸(citrulline)的生成:
乌氨酸氨基甲酰转移酶(ornithinetranscarbamoylase)存在于线粒体中,通常与CPS-I形成酶的复合物催化氨基甲酰磷酸转甲酰基给鸟氨酸生成瓜氨酸。(注意:鸟氨酸,瓜氨酸均非标准α-氨基酸,不出现在蛋白质中)。此反应在线粒体内进行,而鸟氨酸在胞液中生成,所以必需通过一特异的穿棱系统进入线粒体内。

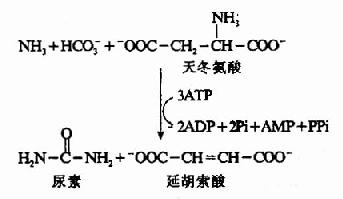
3.精氨酸代琥珀酸(Argininosuccinate)的合成。
瓜氨酸穿过线粒体膜进入胞浆中,在胞浆中由精氨酸代琥珀酸合成酶(Argininosuccinate Synthetase)催化瓜氨酸的脲基与天冬氨酸的氨基缩合生成精氨酸代琥珀酸,获得尿素分子中的第二个氮原子。此反应由ATP供能。
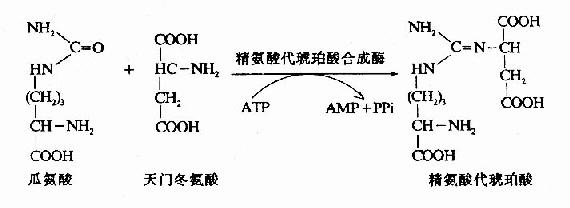
4.精氨酸(Arginine)的生成
精氨酸代琥珀酸裂解酶(Argininosuccinase)催化精氨酸代琥珀酸裂解成精氨酸和延胡索酸
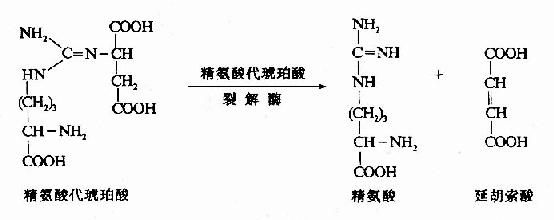
上述反应中生成的延胡索酸可经三羧酸循环的中间步骤生成草酰乙酸,再经谷草转氨酶催化转氨作用重新生成天冬氨酸。由此,通过延胡索酸和天冬氨酸,使三羧酸循环与尿素循环联系起来。
5.尿素的生成
尿素循环的最后一步反应是由精氨酸酶(arginase)催化精氨酸水解生成尿素并再生鸟氨酸,鸟氨酸再进入线粒体参与另一轮循环。
尿素合成是一个耗能的过程,合成1分子尿素需要消耗4个高能磷酸键。(3个ATP水解生成2个ADP,2个Pi,1个AMP和PPi)。从尿素循环底物水平上,能量的消耗大于恢复。由L-谷氨酸脱氢酶催化脱氨和延胡索酸经草酰乙酸再生成天冬氨酸反应中均有NADH的生成。经线粒体再氧化可生成6个ATP(图7-8)。
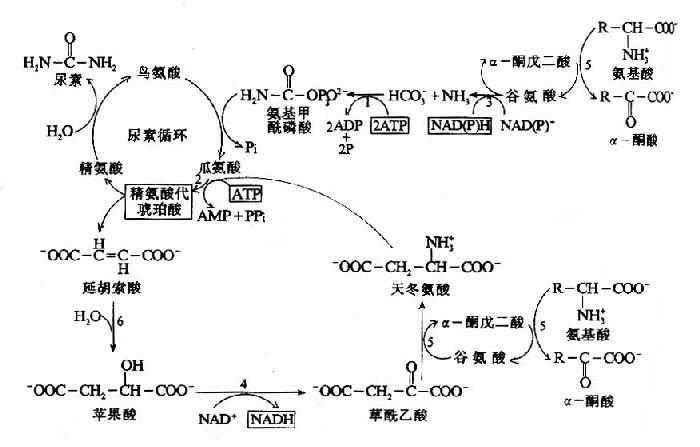
图7-8 尿素循环的能量代谢
6.尿素循环的调节
CPS-I是线粒体内变构酶,其变构激活剂AGA由N?乙酰谷氨酸合成酶催化生成,并由特异水解酶水解。肝脏生成尿素的速度与AGA浓度相关。当氨基酸分解旺盛时,由转氨作用引起谷氨酸浓度升高,增加AGA的合成,从而激活CPS-I,加速氨基甲酰磷酸合成,推动尿素循环。精氨酸是AGA合成酶的激活剂,因此,临床利用精氨酸治疗高氨血症。
(五)高氨血症和氨中毒
正常生理情况下,血氨处于较低水平。尿素循环是维持血氨低浓度的关键。当肝功能严重损伤时,尿素循环发生障碍,血氨浓度升高,称为高氨血症。氨中毒机制尚不清楚。一般认为,氨进入脑组织,可与α酮戊二酸结合成谷氨酸,谷氨酸又与氨进一步结合生成谷氨酰胺,从而使α?酮戊二酸和谷氨酸减少,导致三羧酸循环减弱,从而使脑组织中ATP生成减少。谷氨酸本身为神经递质,且是另一种神经递质γ-氨基丁酸(γ-aminobutyrate,GABA)的前体,其减少亦会影响大脑的正常生理功能,严重时可出现昏迷。
二、α-酮酸的代谢
氨基酸经联合脱氨或其它方式脱氨所生成的α-酮酸有下述去路。
1.生成非必需氨基酸-α-酮酸经联合加氨反应可生成相应的氨基酸。八种必需氨基酸中,除赖氨酸和苏氨酸外其余六种亦可由相应的α-酮酸加氨生成。但和必需氨基酸相对应的α-酮酸不能在体内合成,所以必需氨基酸依赖于食物供应。
2.氧化生成CO2和水这是α-酮酸的重要去路之一。由图7?可以看出α-酮酸通过一定的反应途径先转变成丙酮酸、乙酰CoA、或三羧酸循环的中间产物,再经过三羧酸循环彻底氧化分解。三羧酸循环将氨基酸代谢与糖代谢、脂肪代谢紧密联系起来。
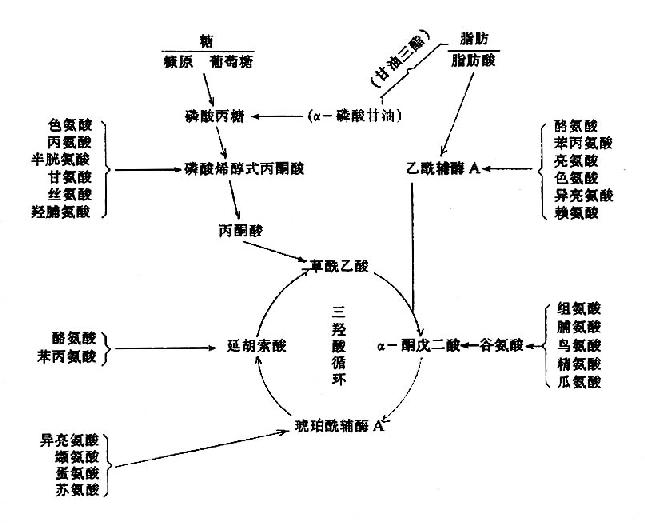
图7-9 氨基酸与糖、脂肪的关系
3.转变生成糖和酮体 使用四氧嘧啶(alloxan)破坏犬的胰岛β-细胞,建立人工糖尿病犬的模型。待其体内糖原和脂肪耗尽后,用某种氨基酸饲养,并检查犬尿中糖与酮体的含量。若饲某种氨基酸后尿中排出葡萄糖增多,称此氨基酸为称生糖氨基酸(glucogenic amino acid);若尿中酮体含量增多,则称为生酮氨基酸(ketogenicamino acid)。尿中二者都增多者称为生糖兼生酮氨基酸(glucogenic and ketogenicamino acid)。从表7-1中可以看出,凡能生成丙酮酸或三羧酸循环的中间产物的氨基酸均为生糖氨基酸;凡能生成乙酰CoA或乙酰乙酸的氨基酸均为生酮氨基酸;凡能生成丙酮酸或三羧酸循环中间产物同时能生成乙酰CoA或乙酰乙酸者为生糖兼生酮氨基酸。
表7-1 氨基酸和糖、脂肪的共有中间代谢产物
| 氨基酸简称 |
共同中间代谢产物 |
生糖或生酮 |
| 天 |
草酰乙酸 |
生糖 |
| 丝、甘、丙、羟、脯、半胱、胱、 |
丙酮酸 |
生糖 |
| 苏 |
丙酮酸、琥珀酰辅酶A |
生糖 |
| 色 |
丙酮酸、乙酰乙酸 |
生糖兼生酮 |
| 谷、组、鸟、精、瓜、脯 |
α-酮戊二酸 |
生糖 |
| 蛋、 缬 |
琥珀酰辅酶A |
生糖 |
| 异亮 |
琥珀酰辅酶A、乙酰辅酶A |
生糖兼生酮 |
| 酪、苯丙 |
乙酰乙酸、延胡索酸 |
生糖兼生酮 |
| 亮 |
乙酰乙酸 |
生酮 |
| 赖 |
乙酰辅酶A、α-酮戊二酸(?) |
生糖兼生酮 |
亮氨酸为生酮氨基酸,赖氨酸、异亮氨酸、色氨酸、苯丙氨酸和酪氨酸为生糖兼生酮氨基酸,其余氨基酸均为生糖氨基酸。
三、脱羧基作用
部分氨基酸可在氨基酸脱羧酶(decarboxylose)催化下进行脱羧基作用(decarboxylation),生成相应的胺,脱羧酶的辅酶为磷酸吡哆醛。
从量上讲,脱羧基作用不是体内氨基酸分解主要方式,但可生成有重要生理功能的胺。下面列举几种氨基酸脱羧产生的重要胺类物质。
1.γ-氨基丁酸(γ-aminobutyric acid GABA)
GABA由谷氨酸脱羧基生成,催化此反应的酶是谷氨酸脱羧酶。此酶在脑、肾组织中活性很高,所以脑中GABA含量较高。
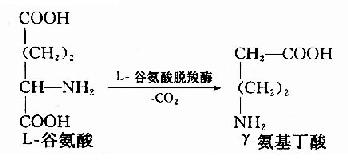
GABA是一种仅见于中枢神经系统的抑制性神经递质,对中枢神经元有普遍性抑制作用。在脊髓,作用于突触前神经末梢,减少兴奋性递质的释放,从而引起突触前抑制,在脑则引起突触后抑制。
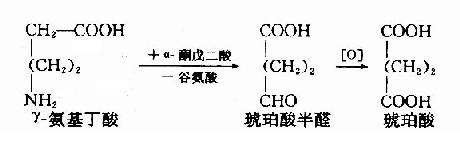
GABA可在GABA转氨酶(GABA-T)作有下与α-酮戊二酸反应生成琥珀酸r-半醛(succinic acid semialdehyde),进而氧化生成琥珀酸。
神经元胞体和突触的线粒体内含有大量的GABA转氨酶。由此就构成了GABA旁路(图7-10)。它能使α?酮戊二酸经此旁路生成琥珀酸,活跃三羧酸循环,可为脑组织提供约20%的能量。谷氨酸具有兴奋作用,GABA有抑制作用,两者可共同调节神经系统的功能。临床上对于惊厥和妊娠呕吐的病人常常使用维生素B6治疗,其机理就在于提高脑组织内谷氨酸脱羧酶的活性,使GABA生成增多,增强中枢抑制作用。
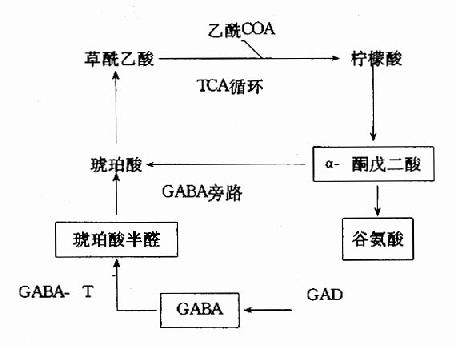
图7-10 脑中TCA循环和GAB代谢旁路
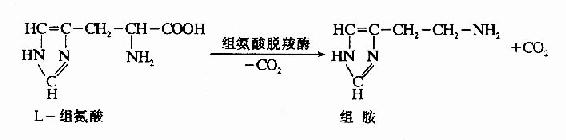
2.组胺(histamine)
由组氨酸脱羧生成。组胺主要由肥大细胞产生并贮存,在乳腺、肺、肝、肌肉及胃粘膜中含量较高。
组胺是一种强烈的血管舒张剂,并能增加毛细血管的通透性。可引起血压下降和局部水肿。组胺的释放与过敏反应症状密切相关。组胺可刺激胃蛋白酶和胃酸的分泌,所以常用它作胃分泌功能的研究。
3.5羟色胺(5?hydroxytryptamine,5?HT)
色氨酸在脑中首先由色氨酸羟化酶(tryoptophanhydroxylase)催化生成5羟色氨酸(5?hydroxy?tryptophan),再经脱羧酶作用生成5羟色胺。
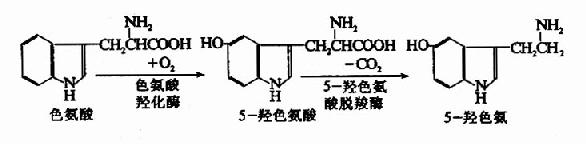
5-羟色胺在神经组织中有重要的功能,目前已肯定中枢神经系统有5-羟色胺能神经元。5-羟色胺可使大部分交感神经节前神经元兴奋,而使付交感节前神经元抑制。
其它组织如小肠、血小板、乳腺细胞中也有5-羟色胺,具有强烈的血管收缩作用。
4.牛磺酸(taurine)
体内牛磺酸主要由半胱氨酸脱羧生成。半胱氨酸先氧化生成磺酸丙氨酸,再由磺酸丙氨酸脱羧酶催化脱去羧基,生成牛磺酸。牛磺酸是结合胆汁酸的重要组成分。

5.多胺(palyamine)
鸟氨酸在鸟氨酸脱羧酶催化下可生成腐胺(putrescine),S-腺苷蛋氨酸(S-adenosyl methionine SAM)在SAM脱羧酶催化脱羧生成S-腺苷-3-甲硫基丙胺。在精脒合成酶(spormidinesynthetase)催化下将S-腺苷-3-甲硫基丙胺的丙基移到腐胺分子上合成精脒(cpermidine),再在精胺合成酶(spermine symthetase)催化下,再将另一分子S-腺苷-3-甲硫基丙胺的丙胺基转移到精脒分子上,最终合成了精胺(sperrmine)。腐胺、精脒和精胺总称为多胺或聚胺polyamine)(图7-11)。
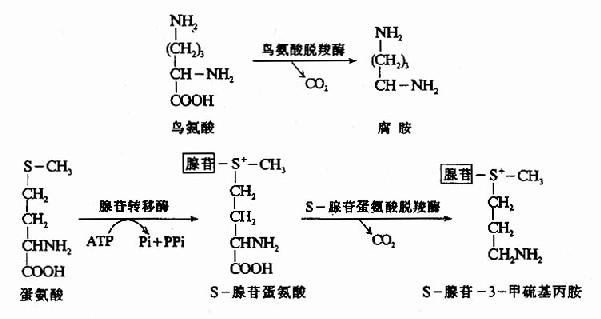
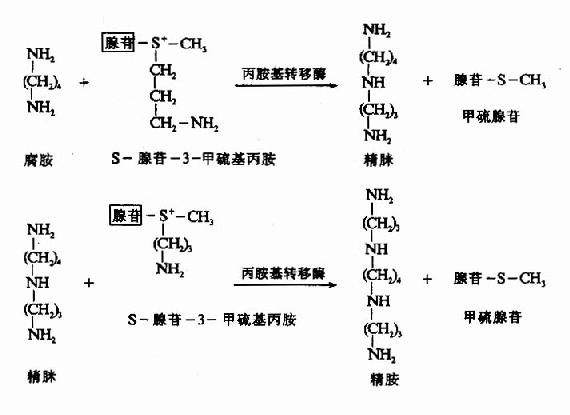
图7-11 多胺的生成
多胺存在于精液及细胞核糖体中,是调节细胞生长的重要物质,多胺分子带有较多正电荷,能与带负电荷的DNA及RNA结合,稳定其结构,促进核酸及蛋白质合成的某些环节。在生长旺盛的组织如胚胎、再生肝及癌组织中,多胺含量升高。所以可将利用血或尿中多胺含量作为肿瘤诊断的辅助指标。
第二节 个别氨基酸代谢
一、一碳单位代谢
某些氨基酸在代谢过程中能生成含一个碳原子的基团,经过转移参与生物合成过程。这些含一个碳原子的基团称为一碳单位(C1unit或one carbon unit)。有关一碳单位生成和转移的代谢称为一碳单位代谢。
体内的一碳单位有:甲基(-CH3,methyl)、甲烯基(-CH2?,methylene),甲炔基(-CH=,methenyl)、甲酰基(-CHO,formyl)及亚氨甲基(-CH=NH,formimino)等。它们可分别来自甘氨酸、组氨酸、丝氨酸、色氨酸、蛋氨酸等(图7-12)。
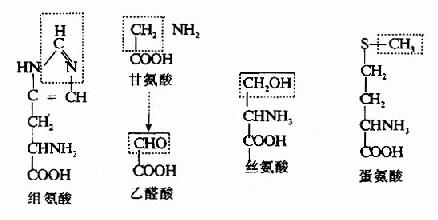
图7-12 一碳单位的来源
(一)一碳单位代酸的辅酶
一碳单位不能游离存在,通常与四氢叶酸(Tetrahydrofolic acid,FH4)结合而转运或参加生物代谢,FH4是一碳单位代谢的辅酶。
四氢叶酸由叶酸(folicacid)衍生而来。叶酸需经二次还原方可转变为活性辅酶形式-FH4(图7-13)。两次还原均由二氢叶酸还原酶(dihyclrofolatereductase)所催化。
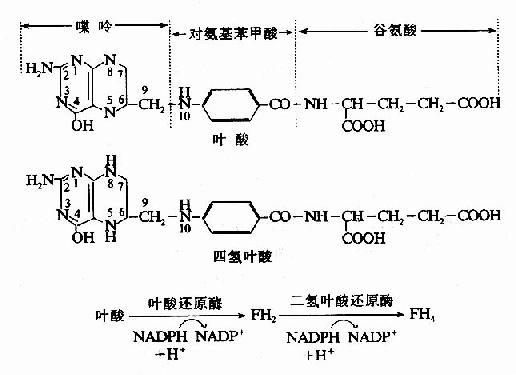
图7-13 四氢叶酸的生成
一碳单位共价连接于FH4分子的N5、N10位或N5和N10位上。
(二)一碳单位的来源及转换
一碳单位主要来源于丝氨酸,在丝氨酸羟甲基转移酶催化为甘氨酸过程中产生的N5,N10?甲烯?FH4;甘氨酸在甘氨酸合成酶(glycine synthase)催化下可分解为CO2,NH+4和N5,N10桟H2桭H4。此外,苏氨酸和丝氨酸都可经相应酶催化转变为丝氨酸。因此亦可产生N5、N10桟H2桭H4。
在组氨酸转变为谷氨酸过程中由亚胺甲基谷氨酸提供了N5桟H=NH?FH4。
色氨酸分解代谢能产生甲酸,甲酸可与FH4结合产生N10桟HO桭H4。
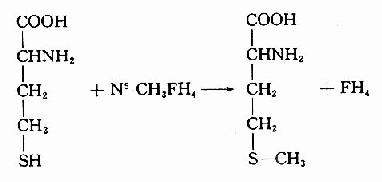
体内一碳单位分别处于甲酸、甲醛不同的氧化水平,在相应的酶促氧化还原反应下可相互转换(图7-14)。这些反应中,N5-CH3-FH4的生成基本是不可逆的。N5-CH3桭H4可将甲基转移给同型半胱氨酸生成蛋氨酸和FH4。催化此反应的酶是N5-CH3FH4同型半胱氨酸甲基转移酶,辅酶为甲基B12。此反应不可逆,故N5-CH3FH4不能自蛋氨酸生成。
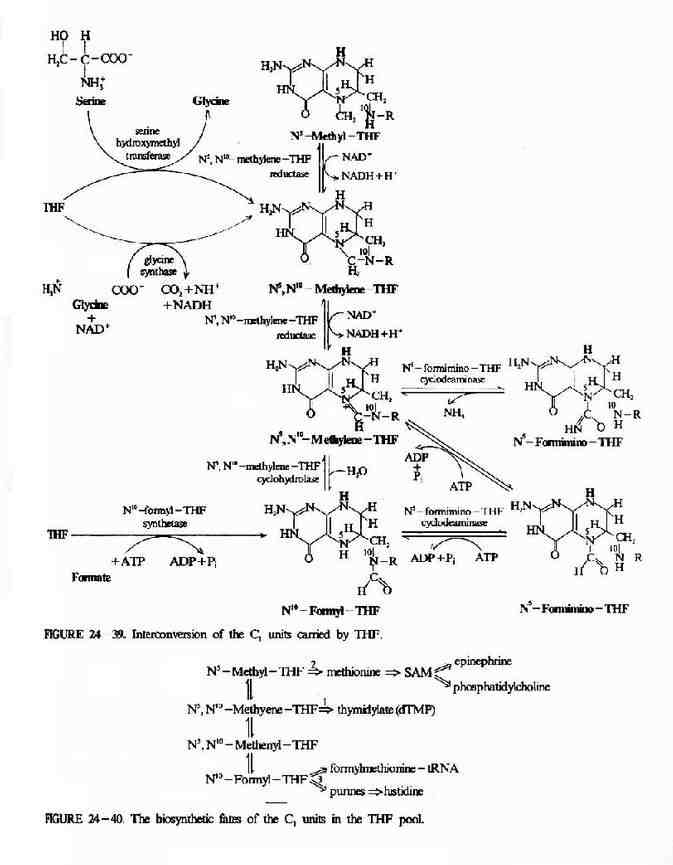
图7-14 一碳单位的相互转化
蛋氨酸分子中的甲基也是一碳单位。在ATP的参与下蛋氨酸转变生成S-腺苷蛋氨酸(Sadenosylmethionine,又称活性蛋氨酸)。S?腺苷蛋氨酸是活泼的甲基供体。因此四氢叶酸并不是一碳单位的唯一载体。
(三)一碳单位的功能
1.一碳单位是合成嘌呤和嘧啶的原料,在核酸生物合成中有重要作用。如N5-N10-CH=FH4直接提供甲基用子脱氧核苷酸dUMP向dTMP的转化。N10-CHO-FH4和N5N10-CH=FH4分别参与嘌呤碱中C2,C3原子的生成。
2.SAM提供甲基可参与体内多种物质合成。例如肾上腺素、胆碱、胆酸等。
一碳单位代谢将氨基酸代谢与核苷酸及一些重要物质的生物合成联系起来。一碳单位代谢的障碍可造成某些病理情况,如巨幼红细胞贫血等。磺胺药及某抗癌药(氨甲喋呤等)正是分别通过干扰细菌及瘤细胞的叶酸、四氢叶酸合成,进而影响核酸合成而发挥药理作用的。
二、含硫氨基酸的代谢
含硫氨基酸共有蛋氨酸、半胱氨酸和胱氨酸三种,蛋氨酸可转变为半胱氨酸和胱氨酸,后两者也可以互变,但后者不能变成蛋氨酸,所以蛋氨酸是必需氨基酸。
(一)蛋氨酸代谢
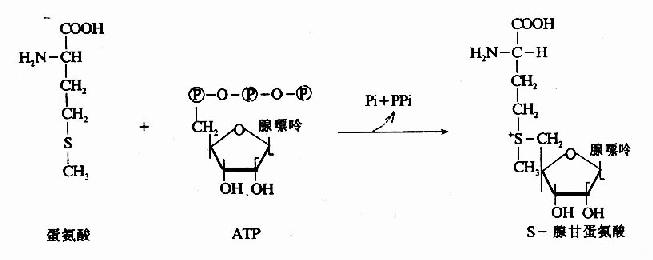
1.转甲基作用与蛋氨酸循环 蛋氨酸中含有S甲基,可参与多种转甲基的反应生成多种含甲基的生理活性物质。在腺苷转移酶催化下与ATP反应生成S-腺苷蛋氨酸(S-adenosglmethiomine,SAM)。SAM中的甲基是高度活化的,称活性甲基,SAM称为活性蛋氨酸。
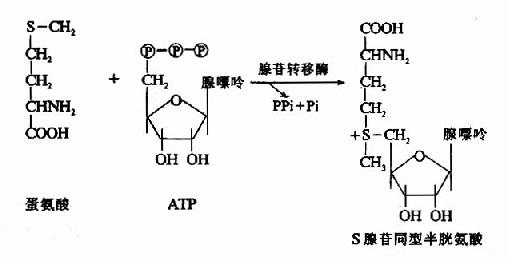
SAM可在不同甲基转移酶(methyl transferase)的催化下,将甲基转移给各种甲接受体而形成许多甲基化合物,如肾上腺素、胆碱、甜菜碱、肉毒碱、肌酸等都是从SAM中获得甲基的。SAM是体内最主要的甲基供体。
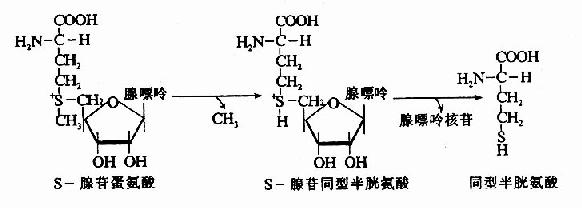
SAM转出甲基后形成S?腺苷同型半胱氨酸S?adenosylhomocystine,SAH),SAH水解释出腺苷变为同型半胱氨酸(homocystine,hCys)。同型半胱氨酸可以接受N5桟H3桯F4提供的甲基再生成蛋氨酸,形成一个循环过程,称为蛋氨酸循环(methionine cycle)。此循环的生理意义在于蛋氨酸分子中甲基可间接通过N5桟H3桭H4由其它非必需氨基酸提供,以防蛋氨酸的大量消耗(图7-15)。
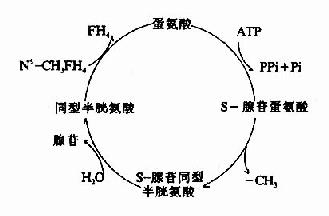
图7-15 S-腺苷蛋氨酸循环
N5-CH3FH4同型半胱氨酸甲基转移酶的辅酶是甲基B12。维生素B12缺乏会引起蛋氨酸循环受阻。临床上可以见到维生素B12缺乏引起的巨幼细胞性贫血。1962年Noronha与Silverman首先提出了甲基陷阱学说(methyl-trap hypothesis),后来Herbert与Zaulsky又作了修改。这个学说认为:由于维生素B12缺乏,引起甲基B12缺乏,使甲基转移酶活性低下,甲基转移反应受阻导致叶酸以N5-CH3FH4形式在体内堆积。这样,其它形式的叶酸大量消耗,以这些叶酸作辅酶的酶活力降低,影响了嘌呤碱和胸腺嘧啶的合成,因而影响核酸的合成,引起巨幼细胞性贫血。也就是说,维生素B12对核酸合成的影响是间接地通过影响叶酸代谢而实现的。
虽蛋氨酸循环可生成蛋氨酸,但体内不能合成同型半胱氨酸,只能由蛋氨酸转变而来,所以体内实际上不能合成蛋氨酸,必须由食物供给。
同型半胱氨酸还可在胱硫醚合成酶(cystathiorinesynthase)催化下与丝氨酸缩合生成胱硫醚(cystathionine),再经胱硫醚酶催化水解生成半胱氨酸,α-酮丁酸和氨。α-酮丁酸转变为琥珀酸单酰CoA,通过三羧酸循环,可以生成葡萄糖、所以蛋氨酸为生糖氨基酸。
2.肌酸的合成 肌酸(creatine)和磷酸肌酸(creatinephosphate)在能量储存及利用中起重要作用。二者互变使体内ATP供应具有后备潜力。肌酸在肝和肾中合成,广泛分布于骨骼肌、心肌、大脑等组织中。肌酸以甘氨酸为骨架,精氨酸提供脒基、SAM供给甲基、在脒基转移酶和甲基转移酶的催化下合成。在肌酸激酶(creatinephosphohinase,CPK)催化下将ATP中桺转移到肌酸分子中形成磷酸肌酸(CP)储备起来(图7-16)。
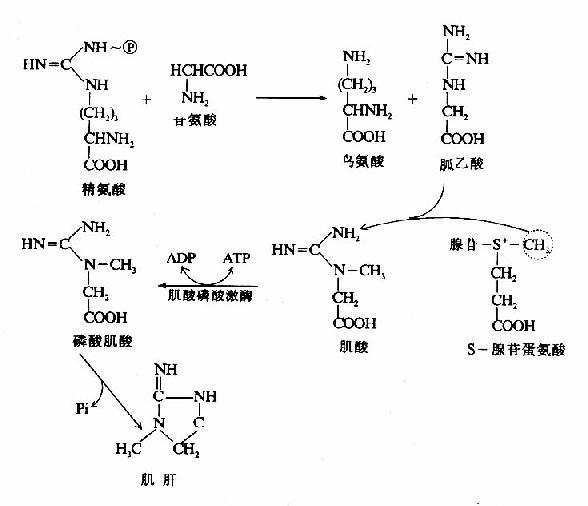
图7-16 肌酸的代谢
CPK由两种亚基组成;即M亚基(肌型)与B亚基(脑型)。有三种同工酶;即MM型(在骨骼肌中)BB型在脑中)和MB型(在心肌中)。心肌梗塞时,血中MB型CPK活性增高,可作辅助诊断的指标之一。
肌酸和磷酸肌酸代谢的终产物是肌酸酐(creatinine)简称肌酐。正常成人,每日尿中肌酐量恒定。肾功能障碍时,检查血或尿中肌酐含量以帮助诊断。
(二)半胱氨酸和胱氨酸的代谢
1.半胱氨酸和胱氨酸的互变 半胱氨酸含巯基(-SH),胱氨酸含有二硫键(S-S-),二者可通过氧化还原而互变。胱氨酸不参与蛋白质的合成,蛋白质中的胱氨酸由半胱氨酸残基氧化脱氢而来。在蛋白质分子中两个半胱氨酸残基间所形成的二硫键对维持蛋白质分子构象起重要作用。而蛋白分子中半胱氨酸的巯基是许多蛋白质或酶的活性基团。
2.半胱氨酸分解代谢 人体中半胱氨酸主要通过两条途径降解为丙酮酸。一是加双氧酶催化的直接氧化途径,或称半胱亚磺酸途径,另一是通过转氨的3-巯基丙酮酸途径。
3.活性硫酸根代谢 含硫氨基酸经分解代谢可生成H2S,H2S氧化成为硫酸。半胱氨酸巯基亦可先氧化生成亚磺基,然后再生成硫酸。其中一部分以无机盐形式从尿中排出,一部分经活化生成3′磷酸腺苷-5'-磷酸硫酸(3'-phosphoadenosine?5'-phosphosulfate,PAPS),即活性硫酸根。
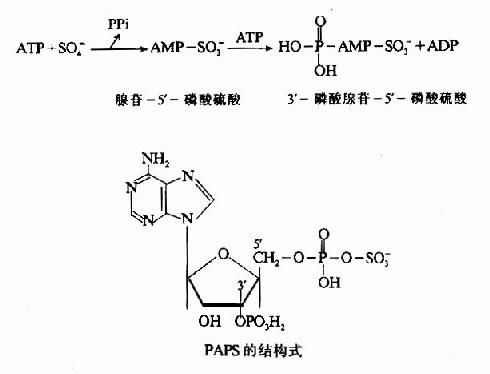
PAPS的性质活泼,在肝脏的生物转化中有重要作用。例如类固醇激素可与PAPS结合成硫酸酯而被灭活,一些外源性酚类亦可形成硫酸酯而增加其溶解性以利于从尿于排出。此外,PAPS也可参与硫酸角质素及硫酸软骨素等分子中硫酸化氨基多糖的合成。
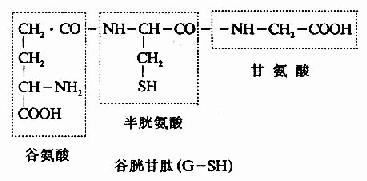
4.谷胱甘肽的合成 谷胱甘肽(glutathiose,r?glutamylcysteinglglycine,GSH)是一种?含γ-酰胺键的三肽,由谷氨酸、半胱氨酸及甘氨酸组成。GSH的合成通过γ-谷氨酰基循环(γ-glutamyl cycle),由Meister提出,又称为Meister循环(图7-17)。γ-谷氨酰基循环有双重作用,一是GSH的再合成,二是通过GSH的合成与分解将外源氨基酸主动转运到细胞内。
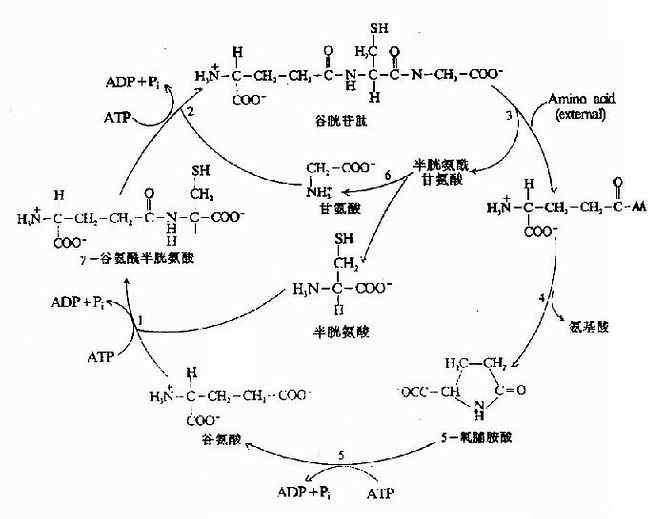
图7-17 γ-谷氨酰基循环
GSH的合成由γ-谷氨酰半胱氨酸合成酶(γ-glutamylcysteinsynthetase)和GSH合成酶(GSHsynthetase)所催化。由ATP水解供能。GSH的分解中γ-谷氨酰转肽酶(γ-glutamyl transpeptidase)、γ-谷氨酰环转移酶(γ-gltamyl cyclotransforase)和5氧脯氨酸酶(5oxoprolinase)及一个细胞内肽酶(protease)所催化。
GSH在人体解毒、氨基酸转运及代谢中均有重要作用。GSH的活性基团是其半胱氨酸残基上的巯基,GSH有氧化型和还原型两种形式,可以互变。
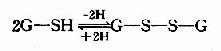
谷胱甘胱还原酶催化上面反应,辅酶为NADPH,细胞中GSH与GSSG的比例为100:1。GSH可保护某些蛋白质及酶分子的巯基不被氧化,从而维持其生物活性。如红细胞中含有较多GSH,对保护红细胞膜完整性及促使高铁血红蛋白还原为血红蛋白均有重要作用。此外,体内产生的过氧化物及自由基,亦可通过含硒的GSH过氧化酶而被清除,如:

三、芳香族氨基酸的代谢
芳香族氨基酸包括苯丙氨酸,酪氨酸和色氨酸,苯丙氨酸和酪氨酸结构相似,在体内苯丙氨酸可转变成酪氨酶,所以合并在一起讨论。
(一)苯丙氨酸和酪氨酸
1.苯丙氨酸在体内一般先转变为酪氨酸。由苯丙氨酸羟化酶(phenylalamine hyolroxylase)催化引入羟基完成,其辅酶为四氢生物嘌呤。反应生成的二氢生物喋呤,由二氢叶酸还原酶催化,借助NADPH+H还原为四氢化合物(图7-18)。
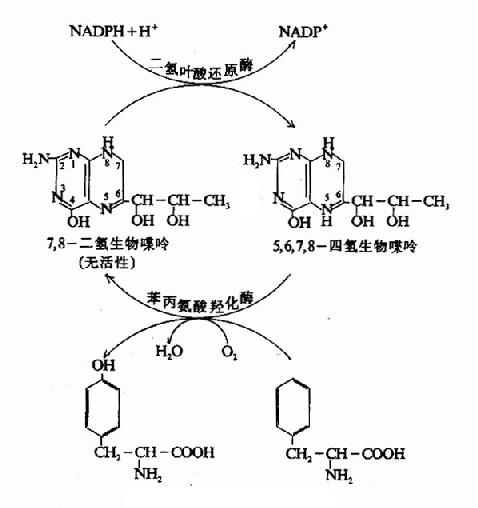
图7-18 酪氨酸的生成
苯丙氨酸羟化酶所催化反应不可逆,体内酷氨酸不能转变为苯丙氨酸。
2.儿茶酚胺与黑色素的合成 酪氨酸经酪氨酸羟化酶(tyrosine hydroxylase)催化生成3,4二羟苯丙氨酸(3,4dihydroxyphenylalanineL-DOPA)(多巴)。此酶也是以四氢生物喋呤为辅酶的加单氧酶,多巴经多巴脱羧酶催化生成多巴胺(dopamine)。多巴胺在多巴胺β-氧化酶(dopamine β?oxidase)催化下使β碳原子羟化,生成去甲肾上腺素(norepinephrine)。而后由SAM提供甲基使去甲肾上腺素甲基化生成肾上腺素(epinephrine)。多巴胺、去甲肾上腺素、肾上腺素统称为儿茶酚胺(catecholamine)。酪氨酸羟化酶是儿茶酚胺合成的限速酶,受终产物的反馈调节(图7-19)。
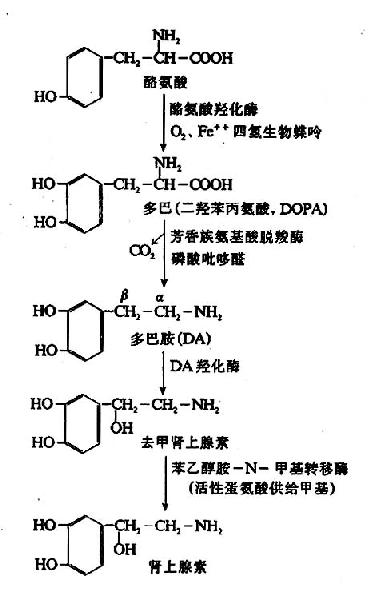
图7-19 儿茶酚胺的合成
在黑色素细胞中,酪氨酸在酪氨酸酶催化下羟化生成多巴,多巴再经氧化生成多巴醌而进入合成黑色素的途径。所形成的多巴醌进一步环化和脱羧生成吲哚醌。黑色素即是吲哚醌的聚合物。人体若缺乏酪氨酸酶,黑色素合成障碍,皮肤、毛发发“白”,称为白化病(albinism)(图7-20)。
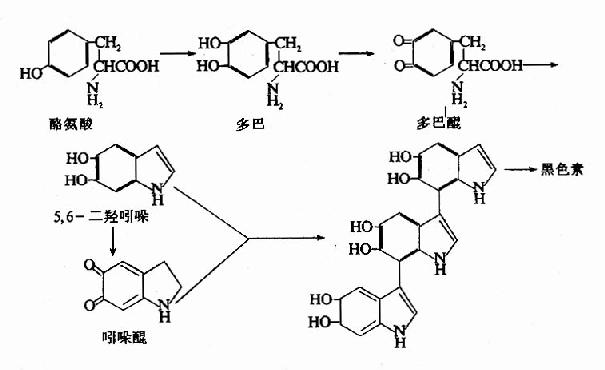
图7-20 黑色素的生成
3.酪氨酸是生糖兼生酮氨基酸 酪氨酸经转氨基作用生成对羟基苯丙酮酸,进一步分解则生成乙酰乙酸和延胡索酸,所以是生糖兼生酮氨基酸。
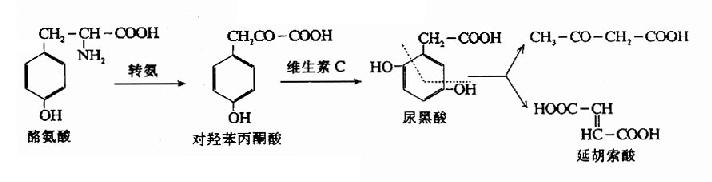
4.代谢障碍 已知在苯丙氨酸和酪氨酸代谢中,有许多代谢性疾患。最重要的是苯丙酮酸尿症(phenylketonuria,PKV),因缺乏苯丙氨酸羟化酶所致。苯丙氨酸不能正常地转变为酪氨酸,体内苯丙氨酸蓄积,并由转氨基作用生成苯丙酮酸(一部分还原为苯乙酸)并从尿液中排出。苯丙酮酸的堆积对中枢神经系统有毒性,故本病伴发智力发育障碍。早期发现时可控制饮食中苯丙氨酸含量,有利于智力发育。
另一代谢疾患为尿黑酸尿症(alkaptonuria)。酪氨酸在分解代谢中生成中间产物尿黑酸,如尿黑酸氧化酶缺乏,则尿黑酸裂环降解受阻,大量尿黑酸排入尿中,经空气氧化为相应的对醌,后者可聚合为黑的色素。此种代谢性疾患一般无严重后果。
此外,巴金森病(Parkinson′sdisease)是由于脑生成多巴胺的功能退化所致的一种严重的神经系统疾病。临床常用L?多巴治疗,L-多巴本身不能通过血?脑屏障无直接疗效,但在相应组织中脱羧可生成多巴胺达到治疗作用。目前,采用将大脑中植移肾上腺髓质,借此生成多巴胺以弥补脑中多巴胺不足,取得较好疗效。
(二)色氨酸的代谢
色氨酸是必需氨基酸。大多数蛋白质中含量均较少,机体对其摄取少,分解亦少。除参加蛋白质合成外,还可经氧化脱羧生成5?羟色胺(前述)。并可降解产生生糖,生酮成分,此过程中产生一碳单位及尼克酸等。
1.色氨酸分解首先在色氨酸-2,3-加双氧酶(tryptophan-2,3-dioxygenase)作用下将吡酪环打开,生成N-甲酰犬尿氨酸(N-Formylkynurenine)。此酶辅基为铁卟啉,Vit C有保护辅基中Fe2+不被氧化的作用,亦可说Vit C是此酶的激活剂。在甲酰化酶(formamidase)的作用下,甲酰犬尿氨酸脱甲酰基生成甲酸和犬尿氨酸,甲酸可参加一碳单位代谢。而犬尿氨酸则有三个不同代谢方向。
(1)犬尿氨酸主要由犬尿氨酸羟化酶(Rynurenine-3-monoxygenase)催化生成3?羟犬尿氨酸(3-hydroxykynurenine),而后由犬尿氨酸酶(kynureninase)(以PLP为辅酶)催化水解裂出丙氨酸,并生成了3-羟邻氨苯甲酸(3-hydroxyanthranilate),丙氨酸可经转氨生成丙酮酸,而3-羟邻苯甲酸经氧化裂环,脱羧等反应生成α?酮乙酸,进而生成乙酰乙酸。因此,色氨酸为生糖兼生酮氨基酸。
(2)少量犬尿氨酸经转氨作用并缩合生成犬尿酸。
(3)少量裂解出丙氨酸后生成邻氨苯甲酸。
2.尼克酸的生成 色氨酸分解代谢中的3-羟邻氨苯甲酸经3-羟邻氨苯丙酸-3,4,-加双氧酶(3-hydroxyanthranilate-3,4-dioxygenase)催化裂环,可生成尼克酸,是构成NAD(P)+的关键成分。这是体内合成维生素的一个特例。
四、支链氨基酸的代谢
支链氨基酸(branched?amino acid,BCAA)包括亮氨酸、异亮氨酸和缬氨酸。三者均为必需氨基酸。分解代谢主要在肌肉组织中进行。它们分属于三类,亮氨酸为生酮氨基酸,缬氨酸为生糖氨基酸,异亮氨酸为生糖兼生酮氨基酸。
三种支链氨基酸分解代谢过程均较复杂,一般可分为二阶段。第一阶段,三种氨基酸前三步反应性质相同,产物类似。均为CoA的衍生物,可称为共同反应阶段。第二阶段则反应各异,经若干步反应,亮氨酸产生乙酰CoA及乙酰乙酰CoA,缬氨酸产生琥珀酸单酰CoA,异亮氨酸产生乙酰CoA及琥珀酸单酰CoA分别纳入生糖或生酮的代谢(图7-21)。
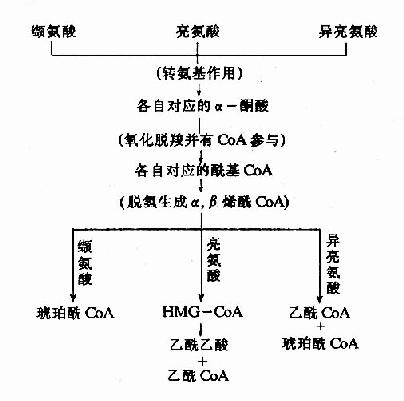
图7-21 支链氨基酸的分解代谢
第三节 氨基酸的生物合成
组成人体蛋白质的氨基酸中,有些氨基酸只能在植物及微生物体内合成,人体必须从食物中摄取,这些氨基酸即必需氨基酸(escential amino acids),其余的氨基酸可利用代谢中间产物合成,称为非必需氨基酸(nonescential amino acids)。(表7-2)除酪氨酸外,体内非必需氨基酸由四种共同代谢中间产物(丙酮酸、草酰乙酸、α-酮戊二酸及3-磷酸甘油)之一作其前体简单合成。如前所述,酪氨酸由苯丙氨酸必需氨基酸羟化生成,严格讲酪氨酸不是非必需氨基酸,对每日膳食中苯丙氨酸的需要量同时亦反映了对酪氨酸的需要量。
表7-2人体中必需和非必需氨基酸
| Essential |
Nonessential |
| Arginine |
Alanine |
| Histidine |
Asparagine |
| Isoleucine |
Aspartate |
| Leucine |
Cysteine |
| Lysine |
Glutamate |
| Methionine |
Glutamine |
| Phenylalanine |
Glycine |
| Threonine |
Proline |
| Tryptophan |
Serine |
| Valine |
Tyrosine |
*Althoughmammals synthesize arginine,they cleave most of it to form urea(Sections 24-2Dand E).
1.丙氨酸,天冬酰胺、天冬氨酸、谷氨酸及谷氨酰胺由丙酮酸、草酰乙酸和α-酮戊二酸合,三种α-酮酸:丙酮酸、草酰乙酸和α-酮戊二酸分别为丙氨酸,天冬氨酸和谷氨酸的前体,经一步转氨反应可生成相应氨基酸(图7-22、反应1-3)。天冬酰胺和谷氨酰胺分别由天冬氨酸和谷胺酸加氨反应生成(图7-22反应4,5)。谷氨酰胺合成酶(glutamine cynthetase)催化谷氨酰胺合成,NH3为氨基供体、反应中消耗ATP生成ADP和Pi。而天冬酰胺由天冬酰胺合成酶(asparagine synthetase)催化合成,利用谷氨酰胺提供氨基、消耗ATP生成AMP+PPi。
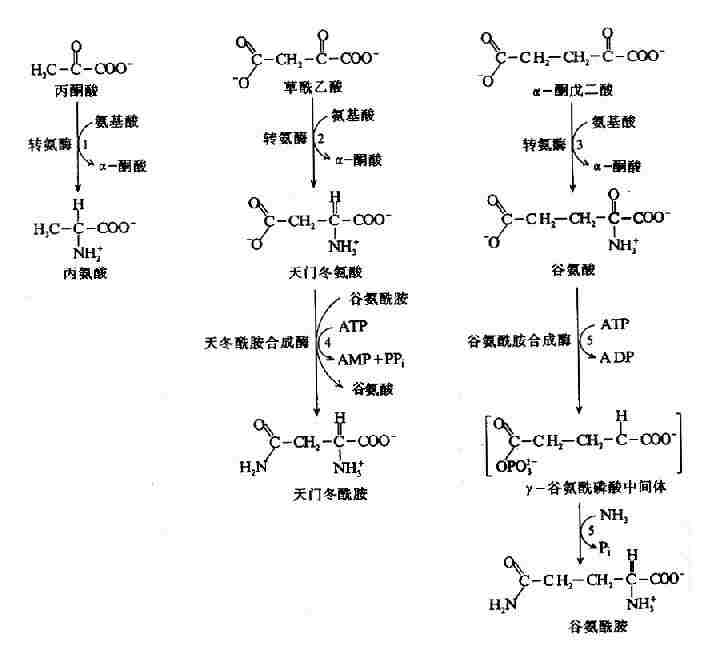
图7-22 氨酸、天冬氨酸、谷氨酸、天冬酰胺和谷氨酰胺的合成
谷氨酰胺是许多生物合成反应的氨基供体,同时也是体内NH2的贮存形式。谷氨酰胺合成酶位于体内氨代谢的中枢位置。实事上,此酶由α-酮戊二酸激活,此种调控作用有利于防止谷氨酸氧化脱氨造成体内氨的堆积。
2.谷氨酸是脯氨酸,鸟氨酸和精氨酸的前体。谷氨酸γ?羧基还原生成醛,继而形成中间Schiff碱,进一步还原可生成脯氨酸(图7?3)。此过程中的中间产物5-谷氨酸半醛(glutamate-5-semialdehyde)在鸟氨酸-δ-氨基转移酶(ornithine-δ-amino-transferase)催化下直接转氨生成鸟氨酸。
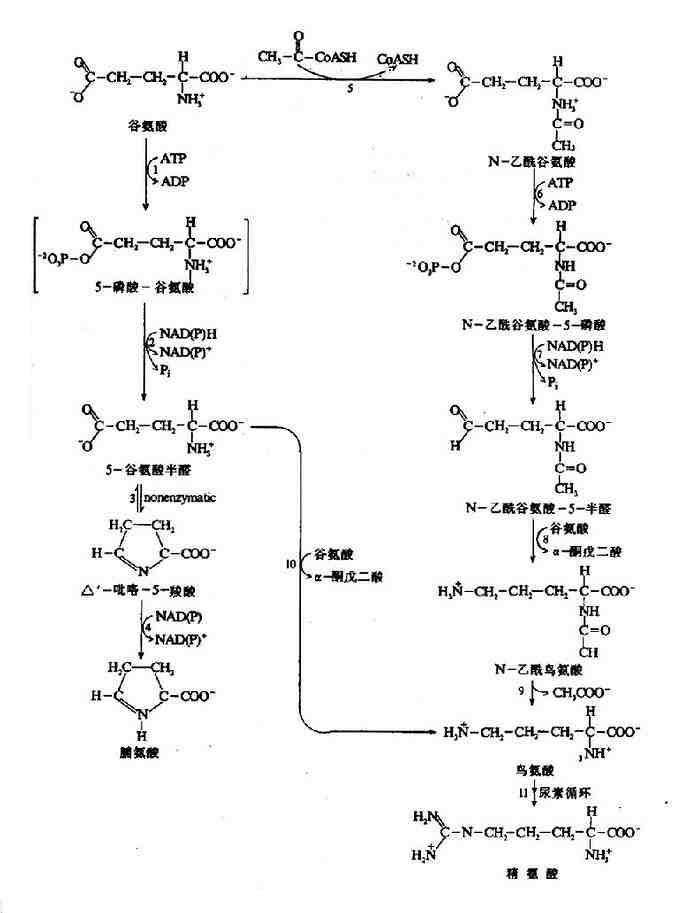
图7-23 由谷氨酸生成脯氨酸、鸟氨酸和精氨酸
3.丝氨酸、半胱氨酸和甘氨酸由三磷酸甘油生成。丝氨酸由糖代谢中间产物3-磷酸甘油经三步反应生成。(1)3-磷酸甘油酸在3-磷酸甘油酸脱氢酶催化下生成了一磷酸羟基丙酮酸(3-phosphohydroxypyruvate)。(2)由谷氨酸提供氨基经转氨作用生成3-磷酸丝氨酸(3-phosphoserine)。(3)3?磷酸丝氨酸水解生成丝氨酸。
丝氨酸以两种途径参与甘氨酸的合成:(1)由丝氨酸羟甲酰转移酶(serine hydroxymethyltransforese)催化直接生成甘氨酸,同时生成N5,N10-甲酰FH4。(2)由N5,N10-CHO-FH4,CO2和NH+4在甘氨酸合成酶(glycineaynthase)催化下缩合生成。
在蛋氨酸代谢中已讨论过,人体中半胱氨酸可由蛋氨酸分解代谢中间产物同型半胱氨酸和丝氨酸合成,半胱氨酸的巯基来源于必需氨基酸-蛋氨酸,故有人将其称为半必需氨基酸(semiessential amino acid)。而在植物及微生物中,半胱氨酸在丝氨酸乙酰转移酶催化下被乙酰基取代生成O-乙酰丝氨酸(O-acetyl serine)。(2)乙酰基被巯基取代生成半胱氨酸。反应中的羟基由PAPS经PAPS还原酶及亚硫酸还原酶(sulfite reductase)催化生成。

 中医世家
中医世家 浦 标 网
浦 标 网 河南大学精品课程
河南大学精品课程 图书资料室
图书资料室