核苷酸在人体内广泛分布,具有多种生物学功能:①核苷酸是构成核酸的基本单位,这是其最主要功能。②储存能量。三磷酸核苷酸,尤其是ATP是细胞的主要能量形式。另外,一些活化的中间产物,如UDP?葡萄糖,亦含有核苷酸成分。③参与代谢和生理调节:许多代谢过程受到体内ATP、ADP或AMP水平的调节。cAMP(或cGMP)是多种细胞膜激素受体的调节作用的第二信使。④组成辅酶。如腺苷酸可作为NAD+、ANDP+、FMN、FAD及CoA等的组成成分。
几乎所有细胞均可以从头合成及补救合成两种途径合成核苷酸。本章重点讨论核苷酸的生物合成过程。同时学习其合成的调节及缺陷,并了解核苷酸的分解代谢等内容。
第一节 核苷酸的化学结构
核苷酸是组成核酸的基本单位。组成DNA的核苷酸是脱氧核糖核苷酸(deoxyribonucleotide)。组成RNA的是核糖核苷酸(ribonudeotide)。核苷酸可以进一步水解为核苷(nucleoside)和磷酸,核苷又可以水解为戊糖(pentose)和碱基(base)(图8-1)

图8-1 核酸的组成
核苷酸中的戊糖有核糖(ribose)和脱氧核糖(deoxyribose)两种,分别存在于核糖核苷酸和脱氧核糖核苷酸中。核苷酸中的碱基均为含氮杂环化合物,分属于嘌呤衍生物和嘧啶衍生物。核苷酸中的嘌呤碱基主要有鸟嘌呤(guanine)和腺嘌呤(adenine);嘧啶碱基主要有:尿嘧啶(uracil)、胞嘧啶(cytosine)和胸腺嘧啶(thymine)。其中尿嘧啶只存在于RNA中,而胸腺嘧啶只存在于DNA中。
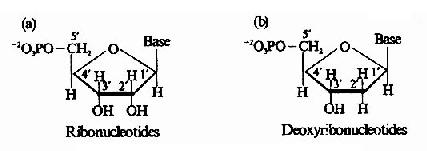
图8-2 核糖核苷酸(a)和脱氧核糖核苷酸(b)的化学结构
碱基与戊糖以糖苷键相连接构成核苷,通常是戊糖的C1′与嘧啶碱的N1或嘌呤碱的N9相连接。核苷中的戊糖与磷酸以磷酸酯键连接构成核苷酸。(图8-2)体内核苷酸大多数是以核糖或脱氧核糖C5′上羟基被磷酸化,形成5′一核苷酸(5′-nucleotide)。除一磷酸核苷外,体内还有核苷的二磷酸酯和三磷酸酯形式。以核糖腺苷酸为例,除AMP外,还有二磷酸腺苷(adenosine?5′-diphosphate ADP)及三磷酸腺苷(adenosine?5′-triphosphate ATP)。二磷酸核苷酸和三磷酸核苷酸多为核苷酸有关代谢中间产物或酶活性及代谢的调节物质。三磷酸核苷酸是参与核酸合成的直接形式,并同时为生理储能和供能的重要形式。
第二节 嘌呤核苷酸的合成代谢
一、嘌呤核苷酸的合成
体内嘌呤核苷酸的合成有两条途径:①利用磷酸核糖、氨基酸、一碳单位及CO2等简单物质为原料合成嘌呤核苷酸的过程,称为从头合成途径(denovo synthesis),是体内的主要合成途径。②利用体内游离嘌呤或嘌呤核苷,经简单反应过程生成嘌呤核苷酸的过程,称重新利用(或补救合成)途径(saluagepathway)。在部分组织如脑、骨髓中只能通过此途径合成核苷酸。
(一)嘌呤核苷酸的从头合成
早在1948年,Buchanan等采用同位素标记不同化合物喂养鸽子,并测定排出的尿酸中标记原子的位置的同位素示踪技术,证实合成嘌呤的前身物为:氨基酸(甘氨酸、天门冬氨酸、和谷氨酰胺)、CO2和一碳单位(N10甲酰FH4,N、N10-甲炔FH4)(图8-3)。
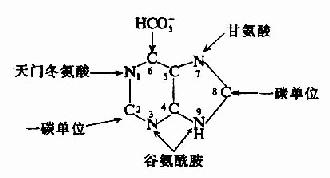
图8-3 嘌呤环合成的原料来源
随后,由Buchanan和Greenberg等进一步搞清了嘌呤核苷酸的合成过程。出人意料的是,体内嘌呤核苷酸的合成并非先合成嘌呤碱基,然后再与核糖及磷酸结合,而是在磷酸核糖的基础上逐步合成嘌呤核苷酸。嘌呤核苷酸的从头合成主要在胞液中进行,可分为两个阶段:首先合成次黄嘌呤核苷酸(inosine monophosphate IMP);然后通过不同途径分别生成AMP和GMP。下面分步介绍嘌呤核苷酸的合成过程。
1.IMP的合成:IMP的合成包括11步反应:(图8-4)
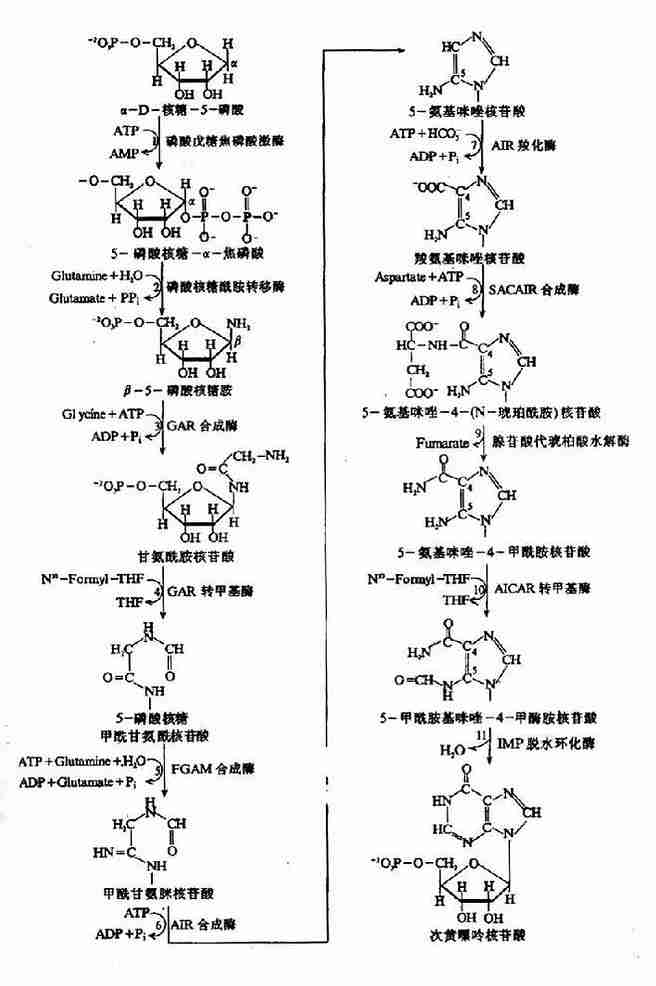
图8-4 IMP的合成
(1)5-磷酸核糖的活化:嘌呤核苷酸合成的起始物为α-D-核糖-5-磷酸,是磷酸戊糖途径代谢产物。嘌呤核苷酸生物合成的第一步是由磷酸戊糖焦磷酸激酶(ribosephosphate pyrophosphohinase)催化,与ATP反应生成5-磷酸核糖-α-焦磷酸(5-phosphorlbosyl?α-pyrophosphatePRPP)。此反应中ATP的焦磷酸根直接转移到5-磷酸核糖C1位上。PRPP同时也是嘧啶核苷酸及组氨酸、色氨酸合成的前体。因此,磷酸戊糖焦磷酸激酶是多种生物合成过程的重要酶,此酶为一变构酶,受多种代谢产物的变构调节。如PPi和2,3-DPG为其变构激活剂。ADP和GDP为变构抑制剂。
(2)获得嘌呤的N9原子:由磷酸核糖酰胺转移酶(amidophosphoribosyltransterase)催化,谷氨酰胺提供酰胺基取代PRPP的焦磷酸基团,形成β-5-磷酸核糖胺(β-5-phosphoribasylamine PRA)。此步反应由焦磷酸的水解供能,是嘌呤合成的限速步骤。酰胺转移酶为限速酶,受嘌呤核苷酸的反馈抑制。
(3)获得嘌呤C4、C5和N7原子:由甘氨酰胺核苷酸合成酶(glycinamide ribotidesynthetase)催化甘氨酸与PRA缩合,生成甘氨酰胺核苷酸(glycinamide ribotide,GAR)。由ATP水解供能。此步反应为可逆反应,是合成过程中唯一可同时获得多个原子的反应。
(4)获得嘌呤C8原子:GAR的自由α-氨基甲酰化生成甲酰甘氨酰胺核苷酸(formylgly?cinamide ribotideFGAR)。由N10-甲酰-FH4提供甲酰基。催化此反应的酶为GAR甲酰转移酶(GAr transtormylase)。
(5)获得嘌呤的N3原子:第二个谷氨酰胺的酰胺基转移到正在生成的嘌呤环上,生成甲酰甘氨脒核苷酸(formylglycinamidine ribotide,FGAM)。此反应为耗能反应,由ATP水解生成ADP+Pi,供能。
(6)嘌呤咪唑环的形成:FGAM经过耗能的分子内重排,环化生成5-氨基咪唑核苷酸(5-aminoimidazole ribotide,AIR)。
(7)获得嘌呤C6原子:C6原子由CO2提供,由AIR羧化酶(AIrcarboxylase)催化生成羧基氨基咪唑核苷酸(carboxyamino imidazoleribotide,CAIR)。
(8)获得N1原子:由天门冬氨酸与AIR缩合反应,生成5-氨基咪唑-4-(N-琥珀酰胺)核苷酸(4-aminoimidazole-4-(N-succinylocarboxamide)ribotide,SACAIR)。此反应与(3)步相似,由ATP水解供能。
(9)去除延胡索酸:SACAIR在SACAIR甲酰转移酶催化下脱去延胡索酸生成5-氨基咪唑-4-甲酰胺核苷酸(5-aminoimidazole-4-carboxamideribotide,AICAR)。(8)、(9)两步反应与尿素循环中精氨酸生成鸟氨酸的反应相似。
(10)获得C2:嘌呤环的最后一个C原子由N10-甲酰-FH4提供,由AICAR甲酰转移酶催化AICAR甲酰化生成5-甲酰胺基咪唑-4-甲酰胺核苷酸(5-formaminoimidazole-4carboxyamideribotide,FAICAR)。
(11)环化生成IMP:FAICAR脱水环化生成 IMP。与反应(6)相反,此环化反应无需ATP供能。
2.由IMP生成AMP和GMP
上述反应生成的IMP并不堆积在细胞内,而是迅速转变为AMP和GMP。AMP与IMP的差别仅是6位酮基被氨基取代(图8-5)。此反应由两步反应完成。(1)天门冬氨酸的氨基与IMP相连生成腺苷酸代琥珀酸(adenylosuccinate),由腺苷酸代琥珀酸合成酶催化,GTP水解供能。(2)在腺苷酸代琥珀酸裂解酶作用下脱去延胡索酸生成AMP。
GMP的生成也由二步反应完成。(1)IMP由IMP脱氢酶催化,以NAD+为受氢体,氧化生成黄嘌呤核苷酸(xanthosinemonophosphate,XMP)。(2)谷氨酰胺提供酰胺基取代XMP中C2上的氧生成GMP,此反应由GMP合成酶催化,由ATP水解供能(图8-5)。
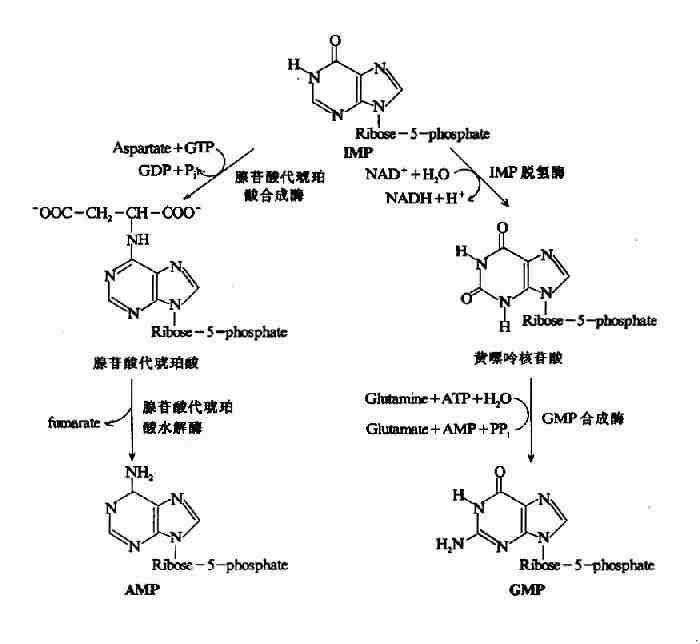
图8-5 IMP分别生成AMP和GMP
3.一磷酸核苷磷酸化生成二磷酸核苷和三磷核苷。
要参与核酸的合成。一磷酸核苷必须先转变为二磷酸核苷再进一步转变为三磷酸核苷。二磷酸核苷由碱基特异的核苷一磷酸激酶(nucleoside monophosphate kinase)催化,由相应一磷酸核苷生成。例如腺苷激酶催化AMP磷酸化生成ADP

二磷酸核苷激酶对底物的碱基及戊糖(核糖或脱氧核糖)均无特异性。此酶催化反应系通过“乒乓机制”,即底物NTP使酶分子的组氨酶残基磷酸化,进而催化底物NDP的磷酸化。反应△G≈0,为可逆反应。
4.嘌呤核苷酸从头合成的调节
从头合成是体内合成嘌呤核苷酸的主要途径。但此过程要消耗氨基酸及ATP。机体对合成速度有着精细的调节。在大多数细胞中,分别调节IMP,ATP和GTP的合成,不仅调节嘌呤核苷酸的总量,而且使ATP和GTP的水平保持相对平衡。嘌呤核苷酸合成调节网可见图8-6。
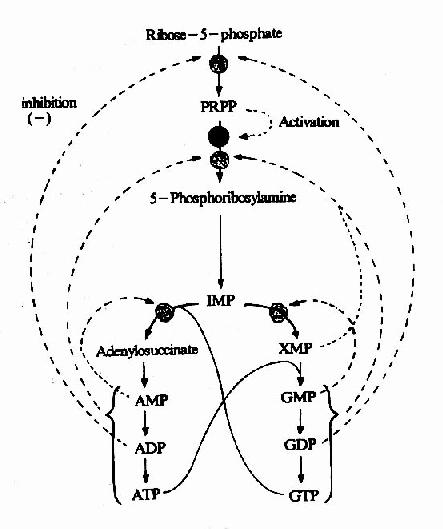
图8-6 嘌呤合成的调节网
IMP途径的调节主要在合成的前二步反应,即催化PRPP和PRA的生成。核糖磷酸焦磷酸激酶受ADP和GDP的反馈抑制。磷酸核糖酰胺转移酶受到ATP、ADP、AMP及GTP、GDP、GMP的反馈抑制。ATP、ADP和AMP结合酶的一个抑制位点,而GTP、GDP和GMP结合另一抑制位点。因此,IMP的生成速率受腺嘌呤和鸟嘌呤核苷酸的独立和协同调节。此外,PRPP可变构激活磷酸核糖酰胺转移酶。
第二水平的调节作用于IMP向AMP和GMP转变过程。GMP反馈抑制IMP向XMP转变,AMP则反馈抑制IMP转变为腺苷酸代琥珀酸,从而防正生成过多AMP和GMP。此外,腺嘌呤和鸟嘌呤的合成是平衡。GTP加速IMP向AMP转变,而ATP则可促进GMP的生成,这样使腺嘌呤和鸟嘌呤核苷酸的水平保持相对平衡,以满足核酸合成的需要。
(二)补救合成途径:
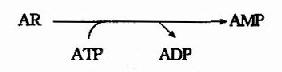
大多数细胞更新其核酸(尤其是RNA)过程中,要分解核酸产生核苷和游离碱基。细胞利用游离碱基或核苷重新合成相应核苷酸的过程称为补救合成(saluage pathway)。与从头合成不同,补救合成过程较简单,消耗能量亦较少。由二种特异性不同的酶参与嘌呤核苷酸的补救合成。腺嘌呤磷酸核糖转移酶(Adenine phosphoribosyl transterase,APRT)催化PRPP与腺嘌呤合成AMP:


人体由嘌呤核苷的补救合成只能通过腺苷激酶催化,使腺嘌呤核苷生成腺嘌呤核苷酸。
嘌呤核苷酸补救合成是一种次要途径。其生理意义一方面在于可以节省能量及减少氨基酸的消耗。另一方面对某些缺乏主要合成途径的组织,如人的白细胞和血小板、脑、骨髓、脾等,具有重要的生理意义。例如Seseh?Nyhan综合征是由于HGPRT的严重遗传缺陷所致。此种疾病是一种性?连锁遗传缺陷,见于男性。患者表现为尿酸增高及神经异常。如脑发育不全、智力低下、攻击和破坏性行为、常咬伤自己的嘴唇、手和足趾,故亦称自毁容貌症。其尿酸增高较易解释,由于HGPRT缺乏,使得分解产生的PRPP不能被利用而堆积,PRPP促进嘌呤的从头合成,从而使嘌呤分解产物-尿酸增高。而神经系统症状的机制尚不清楚。
第三节 嘧啶核苷酸的合成代谢
嘧啶核苷酸合成也有两条途径:即从头合成和补救合成。本节主要论述其从头合成途径。
(一)嘧啶核苷酸的从头合成
与嘌呤合成相比,嘧啶核苷酸的从头合成较简单,同位素示踪证明,构成嘧啶环的N1、C4、C5及C6均由天冬氨酸提供,C3来源于CO2,N3来源于谷氨酰胺。(图8-7)
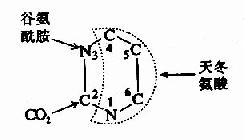
图8-7 嘧啶环合成的原料来源
嘧啶核苷酸的合成是先合成嘧啶环,然后再与磷酸核糖相连而成的。
1.尿嘧啶核苷酸(UMP)的合成,由6步反应完成:(图8-8)
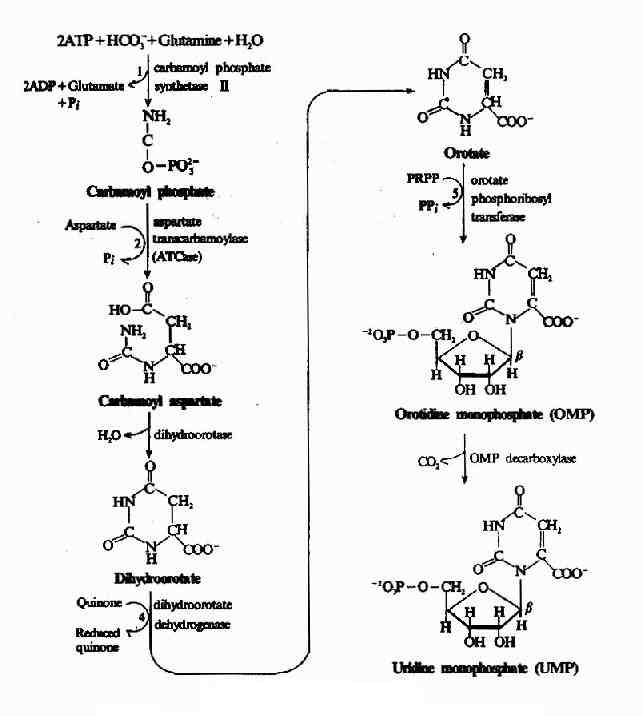
图8-8 UMP的生物合成
(1)合成氨基甲酰磷酸(carbamoyl phosphate):嘧啶合成的第一步是生成氨基甲酰磷酸,由氨基甲酰磷酸合成酶Ⅱ(carbamoyl phosphate synthetase Ⅱ,CPS-Ⅱ)催化CO2与谷氨酰胺的缩合生成。正如氨基酸代谢中所讨论的,氨基甲酰磷酸也是尿素合成的起始原料。但尿素合成中所需氨基甲酰磷酸是在肝线粒体中由CPS-Ⅰ催化合成,以NH3为氮源;而嘧啶合成中的氨基甲酰磷酸在胞液中由CPS?Ⅱ催化生成,利用谷氨酰胺提供氮源。CPS-Ⅰ和CPS-Ⅱ的比较见下表8-1。
(2)合成甲酰天冬氨酸(carbamoyl aspartate):由天冬氨酸氨基甲酰转移酶(aspartate transcarbamoylase,ATCase)催化天冬氨酸与氨基甲酰磷酸缩合,生成氨基甲酰天冬氨酸(carbamoyl aspartate)。此反应为嘧啶合成的限速步骤。ATCase是限速酶,受产物的反馈抑制。不消耗ATP,由氨基甲酰磷酸水解供能。
表8-1 两种氨基甲酰磷酸合成酶的比较
|
氨基甲酰磷酸合成酶Ⅰ |
氨基甲酰磷酸合成酶Ⅱ |
| 分布 |
线粒体现肝脏) |
胞液(所有细胞) |
| 氮源 |
氨 |
谷氨酰胺 |
| 变构激活剂 |
N乙酰谷氨酸 |
无 |
| 反馈抑制剂 |
无 |
UMP(哺乳类动物) |
| 功能 |
尿素合成 |
嘧啶合成 |
(3)闭环生成二氢乳清酸(dihydroortate):由二氢乳清酸酶(dihyolroorotase)催化氨基甲酰天冬氨酸脱水、分子内重排形成具有嘧啶环的二氢乳清酸。
(4)二氢乳清酸的氧化:由二氢乳清酸还原酶(dihydroorotate dehyolrogenase)催化,二氢乳清酸氧化生成乳清酸(orotate)。此酶需FMN和非血红素Fe2+,位于线粒体内膜的外侧面,由醌类(quinones)提供氧化能力,嘧啶合成中的其余5种酶均存在于胞液中。
(5)获得磷酸核糖:由乳清酸磷酸核糖转移酶催化乳清酸与PRPP反应,生成乳清酸核苷酸(orotidine-5′-monophosphate,OMP)。由PRPP水解供能。
(6)脱羧生成UMP:由OMP脱羧酶(omp decarboxylase)催化OMP脱羧生成UMP。
Jones等研究表明,在动物体内催化上述嘧啶合成的前三个酶,即CPS-Ⅱ,天冬氨酸氨基甲酰转移酶和二氢乳清酸酶,位于分子量约210kD的同一多肽链上,是一个多功能酶;因此更有利于以均匀的速度参与嘧啶核苷酸的合成。与此相类似,反应(5)和(6)的酶(乳清酸磷酸核糖转移酶和OMP脱羧酶)也位于同一条多肽链上。嘌呤核苷酸合成的反应(3)、(4)、(6),反应(7)和(8)及反应(10)和(11)也均为多功能酶。这些多功能酶的中间产物并不释放到介质中,而在连续的酶间移动,这种机制能加速多步反应的总速度,同时防止细胞中其它酶的破坏。
2.UTP和CTP的合成
三磷酸尿苷(UTP)的合成与三磷酸嘌呤核苷的合成相似。
三磷酸胞苷(CTP)由CTP合成酶(CTP synthetase)催化UTP加氨生成。(图8-9)动物体内,氨基由谷氨酰胺提供,在细菌则直接由NH3提供。此反应消耗1分子ATP。
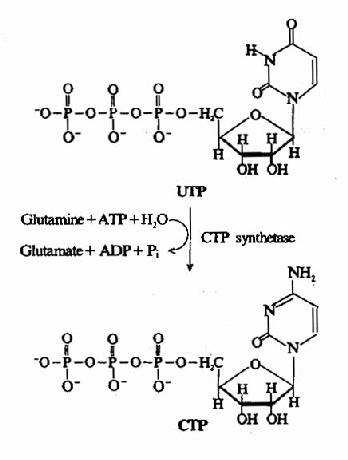
图8-9 由UTP合成CTP
3.嘧啶核苷酸从头合成的调节
在细菌中,天冬氨酸氨基甲酰转移酶(ATCase)是嘧啶核苷酸从头合成的主要调节酶。在大肠杆菌中,ATCase受ATP的变构激活,而CTP为其变构抑制剂。而在许多细菌中、UTP是ATCase的主要变构抑制剂。
在动物细胞中,ATCase不是调节酶。嘧啶核苷酸合成主要由CPS-Ⅱ调控。UDP和UTP抑制其活性,而ATP和PRPP为其激活剂。第二水平的调节是OMP脱羧酶,UMP和CMP为其竞争抑制剂。(图8-10)
此外,OMP的生成受PRPP的影响。
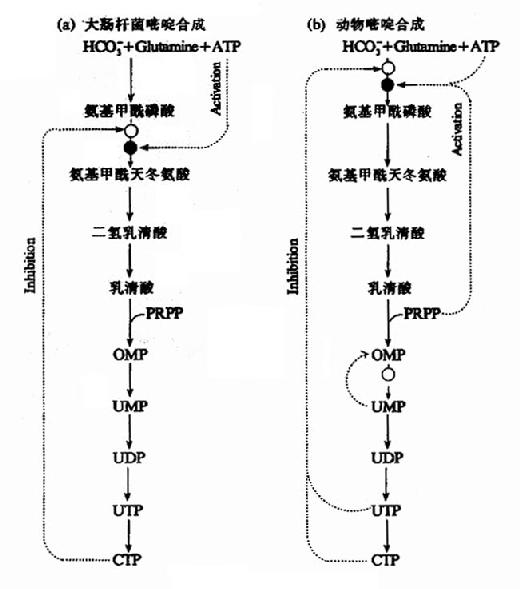
图8-10 嘧啶合成的调节网
4.乳清酸尿症(Orotic aciduria)
乳清酸尿症是一种遗传性疾病,主要表现为尿中排出大量乳清酸、生长迟缓和重度贫血。是由于催化嘧啶核苷酸从头合成反应(5)和(6)的双功能酶的缺陷所致。临床用尿嘧啶或胞嘧啶治疗。尿嘧啶经磷酸化可生成UMP,抑制CPS?Ⅱ活性,从而抑制嘧啶核苷酸的从头合成。
第四节 脱氧核糖核苷酸的生成
DNA与RNA有两方面不同:(1)其核苷酸中戊糖为2?脱氧核糖而非核糖。(2)含有胸腺嘧啶碱基,不含尿嘧啶碱基。
图8-11 大肠杆菌硫氧化还原
蛋白的320残基亚单位结构图
(一)脱氧核糖的生成:
脱氧核糖核苷酸是通过相应核糖核苷酸还原,以H取代其核糖分子中C2上的羟基而生成,而非从脱氧核糖从头合成。此还原作用是在二磷酸核苷酸(NDP)水平上进行的。(此处N代表A、G、U、C等碱基)。
催化脱氧核糖核苷酸生成的酶是核糖核苷酸还原酶(ribonudeotide reductase)。已发现有三种不同的核糖核苷酸还原酶,此反应过程较复杂。核糖核苷酸还原酶催化循环反应的最后一步是酶分子中的二硫键还原为具还原活性的巯基的酶再生过程。硫氧化还原蛋白(thioredoxin)是此酶的一种生理还原剂,由108个氨基酸组成,分子量约12kD。含有一对邻近的半胱氨酸残基(图8?11)。所含硫基在核糖核苷酸还原酶作用下氧化为二硫键,后者再在在硫氧化还原蛋白还原酶(thioredoxinreductase)催化,由NADPH供氢重新还原为还原型的硫氧化还原蛋白。因此,NADPH是NDP还原为dNDP的最终还原剂。(图8-12)
核糖核苷酸还原酶是一种变构酶,包括B1、B2两个亚基,只有B1与B2结合时。才具有酶活性。在DNA合成旺盛、分裂速度快的细胞中,核糖核苷酸还原酶系活性较强。
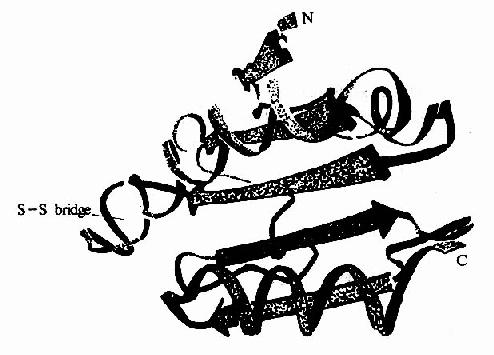
图8-12 大肠杆菌硫氧化还原蛋白的X线衍射结构
(二)脱氧核糖核苷酸合成的调节
四种dNTP的合成水平受到反馈调节,同时保持dNTP的适当比例也是细胞正常生长所必需的。实际上,缺少任一种dNTP都是致命的,而一种dNTP过多也可致突变,因为过多的dNTP可错误掺入DNA链中。核糖核苷酸还原酶的活性对脱氧核糖核苷酸的水平起着决定作用。各种dNTP通过变构效应调节不同脱氧核糖核苷酸生成。因为,某一种特定NDP经还原酶作用生成dNDP时,需要特定NTP的促进,同时受到另一些NTP的抑制(表8?2)。通过调节使4种dNTP保持适当的比例。
表8-2 核糖核苷酸还原酶的别构调节
| 作用物 |
主要促进剂 |
主要抑制剂 |
| CDP |
ATP |
dATP、dGTP、dTTP |
| UDP |
ATP |
dATP、dGTP |
| ADP |
dGTP |
dATP、ATP |
| GDP |
dTTP |
dATP |
例如,当存在混合的NDP底物时,由ATP促使CDP和UDP还原生成dUDP和dCDP。经dUDP转变为dTTP(后述),dTTP则反馈抑制CDP和UDP还原,同时促进dGDP的生成,dGDP磷酸化生成dGTP则抑制GDP、CDP和UDP的还原,而促进ADP的还原生成dADP。当dATP升高与酶
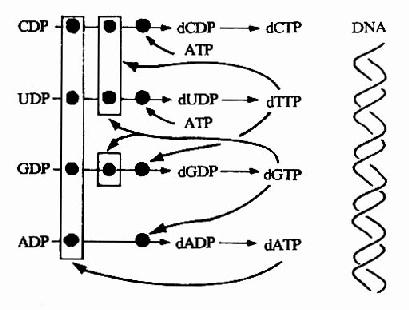
图8-13 脱氧核苷酸合成调节网络
活性位点结合,则抑制所有NDP的还原反应(图8-13)。细胞内dCTP和dTTP的适当比例并非由核糖核苷酸还原酶调节,而是通过脱氧胞嘧啶脱氨酶(deoxycytidinedeaminase)决定。此酶催化dUMP的生成,dUMP则是dTTP的前体。此酶受dCTP激活,受dTTP抑制。
dNTP由dNDP磷酸化生成:
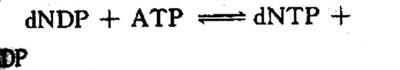
由二磷酸核苷酸激酶(mucleosidediphosphafe kinase),催化与催化NDP磷酸化的反应相似。
(三)脱氧胸腺嘧啶核苷酸的生成
脱氧胸腺嘧啶核苷酸(dTMP)是由脱氧尿嘧啶核苷酸(dUMP)甲基化生成。而dUMP由dUTP水解生成:
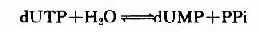
体内进行此种“浪废”能量的反应过程的意义在于:细胞必须减少细胞内dUTP浓度以防止脱氧尿嘧啶掺入DNA中,因为合成DNA的酶系不能有效识别dUTP和dTTP。
dUMP甲基化生成dTMP由胸腺嘧啶合成酶(thymidylatesynthetase,TS)催化,N5,N10-甲烯FH4提供甲基(图8-14)。N5、N10-甲烯-FH4提供甲基后生成的FH2又可以再经二氢叶酸还原酶的作,重新生成四氢叶酸。
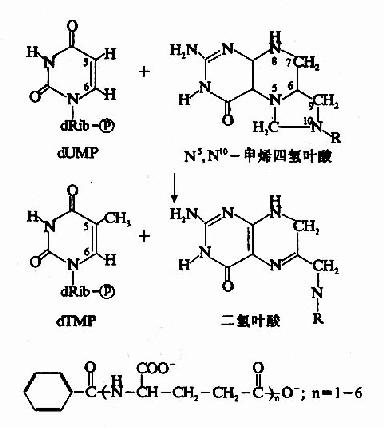
图8-14 dTMP的生成
第五节 核苷酸的分解代谢
食物中的核酸多与蛋白质结合为核蛋白,在胃中受胃酸的作用,或在小肠中受蛋白酶作用,分解为核酸和蛋白质。核酸主要在十二指肠由胰核酸酶(pancreatic nucleases)和小肠磷酸二酯酶(phosphodiesterases)降解为单核苷酸。核苷酸由不同的碱基特异性核苷酸酶(nucleotidases)和非特异性磷酸酶(phosphatases)催化,水解为核苷和磷酸。核苷可直接被小肠粘膜吸收,或在核苷酶(nucleosidases)和核苷磷酸化酶(nucleoside phosphorylases)作用下,水解为碱基,戊糖或1-磷酸戊糖:
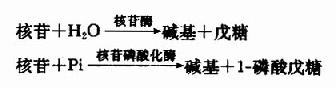
体内核苷酸的分解代谢与食物中核苷酸的消化过程类似,可降解生成相应的碱基,戊糖或1-磷酸核糖。1-磷酸核糖在磷酸核糖变位酶催化下转变为5-磷酸核糖,成为合成PRPP的原料。碱基可参加补救合成途径,亦可进一步分解。
一、嘌呤的分解代谢
嘌呤核苷酸可以在核苷酸酶的催化下,脱去磷酸成为嘌呤核苷,嘌呤核苷在嘌呤核苷磷酸化酶(purine nucleoside phosphorylase,PNP)的催化下转变为嘌呤。嘌呤核苷及嘌呤又可经水解,脱氨及氧化作用生成尿酸(图8-15)
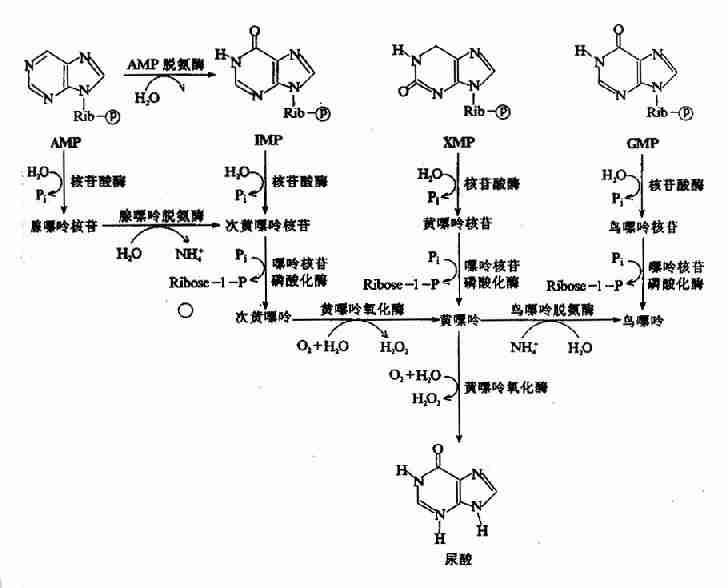
图8-15 嘌呤核苷酸的分解代谢
哺乳动物中,腺苷和脱氧腺苷不能由PNP分解,而是在核苷和核苷酸水平上分别由腺苷脱氨酶(adenosine deaminase,ADA)和腺苷酸脱氨酸(AMPdeaminase)催化脱氨生成次黄嘌呤核苷或次黄嘌呤核苷酸。它们再水解成次黄嘌呤,并在黄嘌呤氧化酶(xanthineoxidase)的催化下逐步氧化为黄嘌呤和尿酸(uric acid)。ADA的遗传性缺乏,可选择性清除淋巴细胞,导致严重联合免疫缺陷病(Severecombined immunodeficiency olisease,SCID)。
体内嘌呤核苷酸的分解代谢主要在肝脏、小肠及肾脏中进行。正常生理情况下,嘌呤合成与分解处于相对平衡状态,所以尿酸的生成与排泄也较恒定。正常人血浆中尿酸含量约0.12~0.36mmol/L(2~6mg/dl)。男性平均为0.27mmol/L(4.5mg/dl),女性平均为0.21mmol/L(3.5mg/dl)左右。当体内核酸大量分解(白血病、恶性肿瘤等)或食入高嘌呤食物时,血中尿酸水平升高,当超过0.48mmol/L(8mg/dl)时,尿酸盐将过饱合而形成结晶,沉积于关节、软组织、软骨及肾等处,而导致关节炎、尿路结石及肾疾患,称为痛风症。痛风症多见于成年男性,其发病机理尚未阐明。临床上常用别嘌呤醇(allopurinol)治疗痛风症。别嘌呤醇与次黄嘌呤结构类似,只是分子中N8,与C2互换了位置,故可抑制黄嘌呤氧化酶,从而抑制尿酸的生成。同时,别嘌呤在体内经代谢转变,与PRPP生成别嘌呤核苷酸,不仅消耗了PRPP,使其含量下降,而且还能反馈抑制PRPP酰胺转移酶,阻断嘌呤核苷酸的从头合成。
二、嘧啶核苷酸的分解代谢

嘧啶核苷酸的分解代谢途径与嘌呤核苷酸相似。首先通过核苷酸酶及核苷磷酸化酶的作用,分别除去磷酸和核糖,产生的嘧啶碱再进一步分解。嘧啶的分解代谢主要在肝脏中进行。分解代谢过程中有脱氨基、氧化、还原及脱羧基等反应。胞嘧啶脱氨基转变为尿嘧啶。尿嘧啶和胸腺嘧啶先在二氢嘧啶脱氢酶的催化下,由NADPH+H+供氢,分别还原为二氢尿嘧啶和二氢胸腺嘧啶。二氢嘧啶酶催化嘧啶环水解,分别生成β-丙氨酸(β-alanine)和β-氨基异丁酸(β-aminosiobutyrate)。β-丙氨酸和β氨基异丁酸可继续分解代谢。β-氨基异丁酸亦可随尿排出体外。食入含DNA丰富的食物、经放射线治疗或化学治疗的患者,以及白血病患者,尿中β-氨基异丁酸排出量增多。嘧啶核苷酸分解代谢见图8-16。
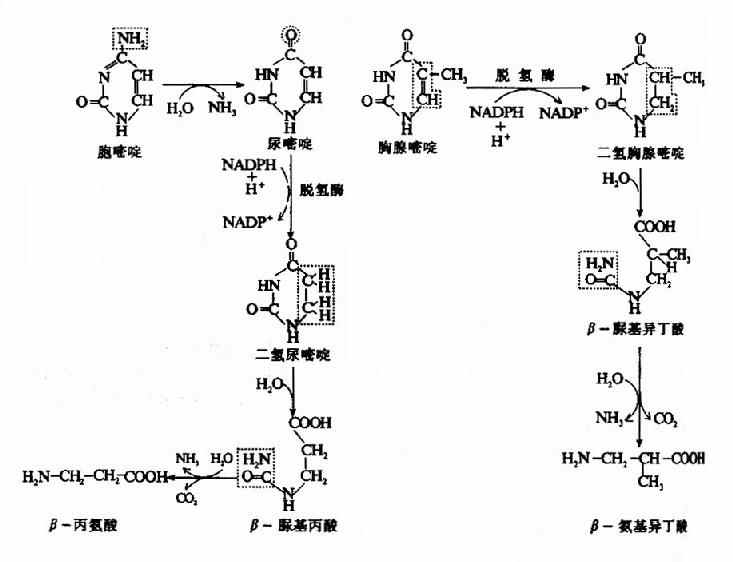
图8-16 嘧啶核苷酸的分解代谢

 中医世家
中医世家 浦 标 网
浦 标 网 河南大学精品课程
河南大学精品课程 图书资料室
图书资料室