第七章索引
第一节 能量代谢
新陈代谢是机体生命活动的基本特征,新陈代谢包括物质代谢与相传伴的能量代谢,简称代谢。
糖、脂肪、蛋白质三种营养物质,经消化转变成为可吸收的小分子营养物质而被吸收入血。在细胞中,这些营养物质经过同化作用(合成代谢),构筑机体的组成成分或更新衰老的组织;同时经过异化作用(分解代谢)分解为代谢产物。合成代谢和分解代谢是物质代谢过程中互相联系的、不可分割的两个侧面。
在分解代谢过程中,营养物质蕴藏的化学能便释放出来。这些化学能经过转化,便成了机体各种生命活动的能源,所以说分解是代谢的放能反应。而在合成代谢过程中,需要供给能量,因此是吸能反应。可见,在物质代谢过程中,物质的变化与能量的代谢是紧密联系着的。生物体内物质代谢过程中所伴随的能量释放、转移和利用等,称为能量代谢(energy metabolism)。
机体所需的能量来源于食物中的糖、脂肪和蛋白质。这些能源物质分子结构中的碳氢键蕴藏着化学能,在氧化过程中碳氢键断裂,生成CO2和 H2O,同时释放出蕴藏的能。这些能量的50%以上迅速转化为热能,用于维持体温,并向体外散发。其余不足50%则以高能磷酸键的形式贮存于体内,供机体利用。体内最主要的高能磷酸键化学物是三磷酸腺苷(ATP)。此外,还可有高能硫酯键等。机体利用ATP去合成各种细胞组成分子、各种生物活性物质和其他一些物质;细胞利用ATP去进行各种离子和其它一些物质的主动转运,维持细胞两侧离子浓度差所形成的势能;肌肉还可利用ATP所载荷的自由能进行收缩和舒张,完成多种机械功。总的看来,除骨骼肌运动时所完成的机械功(外功)以外,其余的能量最后都转变为热能。例如心肌收缩所产生的势能(动脉血压)与动能(血液流速),均于血液在血管内流动过程中,因克服血流内、外所产生的阻力而转化为热能。在人体内,热能是最“低级”形式的能,热能不能转化为其它形式的能,不能用来作功。
本节主要叙述整个机体的能量代谢测定的原理与方法,基础代谢以及机体在某些状态下的代谢等问题,不涉及能量代谢的各个方面。
通用的法定计量的热量单位为焦耳(Joules.J)。过去热量单位是卡或千卡,1卡=4.187焦耳,1千卡=4.187千焦耳(kJ)。1焦耳/s为1瓦特。
一、能量代谢测定的原理和方法
热力学第一定律指出:能量由一种形式转化为另一种形式的过程中,既不能增加,也不减少。这是所有形式的能量(动能、热能、电能入化学能)互相转化的一般规律,也就是能量守恒定律。机体的能量代谢也遵循这一规律,即在整个能量转化过程中,机体所利用的蕴藏于食物中的化学能与最终转化成的热能和所作的外功,按能量来折算是完全相等的。因此,测定在一定时间内机体所消耗的食物,或者测定机体所产生的热量与所做的外功,都可测算出整个机体的能量代谢率(单位时间内所消耗的能量)。
测定整个机体单位时间内发散的总热量,通常有两类方法:直接测热法和间接测热法。
(一)直接测热法
直接测热法(direct calormetry)是测定整个机体在单位时间内向外界环境发散的总热量。此总热量就是能量代谢率。如果在测定时间内做一定的外功,应将外功(机械功)折算为热量一并计入。图7-1是本世纪初Arwater-Benedict所设计的呼吸热量计的结构模式图。它是由隔热密封的房间,其中设一个铜制的受试者居室。用调节温度的装置控制隔热壁与居室之间空气的温度,使之与居室内的温度相等,以防居室内的热量因传导而丧失。这样,受试者机体所散发的大部分热量便被居室内管道中流动的水所吸收。根据流过管道的水量和温度差,将水的比热考虑在内,就可测出水所吸收的热量。当然,受试者发散的热量有一部分包含在不感蒸发(参看第二节)量中,这在计算时也要加进去。受试者呼吸的空气由进出居室的气泵管道系统来供给。此系统中装有硫酸和钠石灰,用业吸收水蒸气和CO2。管道系统中空气中的O2则由氧气筒定时补给。
直接测热法的设备复杂,操作繁锁,使用不便,因而极少应用。一般都采用间接测热法。
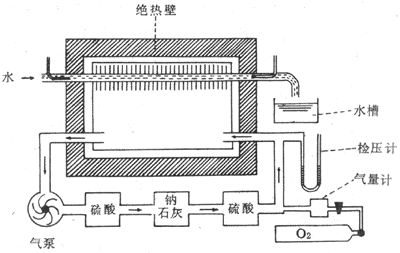
图7-1 直接测热装置示意图
(二)间接测热法
在一般化学反应中,反应物的量与产物量之间呈一定的比例关系,这就是定比定律。例如,氧化1mol葡萄糖,需要6mol氧,同时产生6molCO2和6molH2O,并释放一定量的能。下列反应式表明了这种关系:
C6H12O6+602→6CO2+6H20+△H
同一种化学反应,不论经过什么样的中间步骤,也不论反应条件差异多大,这种定比关系仍然不变。例如,在人本内氧化1mol葡萄糖,同在体外氧化燃烧1mol葡萄糖一样,都要消耗6molCO2和6molH20,而且产生的热量也相等。一般化学反应的这种基本规律也见于人体内营养物质氧化供能的反应(蛋白质的情况下有些出入,参看下文),所以它成了能量代谢间接测热法的重要依据。
间接测热法(indirectcalorimetry)的基本原理就是利用这种定比关系,查出一定时间内整个人体中氧化分解的糖、脂肪、蛋白质各有多少,然后据此计算出该段时间内整个机体所释放出来的热量。因此,必须解决两个问题:一是每种营养物质氧化分解时产生的能量有多少(即食物的热价);二要分清三种营养物质各氧化了多少。
食物的热价应用弹式热量计,在体外测定了一定量的的糖、脂肪和蛋白质燃烧时所释放的热量,并同这三类物质在动物体内氧化到最终产物C02和水时所产生的热量相比较,证明了糖和脂肪在体外燃烧与在体内氧化分解所产生的热量是相等的。于是将1g食物氧化(或在体外燃烧)时所释放出来的能量称为食物的热价(thermalequivalent of food)。食物的热价分为物理热价和生物热价。前者指食物在体外燃烧时释放的热量,后者系食物经过生物氧化所产生的热量。糖(或脂肪)的物理热价和生物热价是相等的,而蛋白质的生物热价则小于它的物理热价。因为蛋白质在体内不能被彻底氧化分解,它有一部分主要以尿素的形式从尿中排泄的缘故。三种营养物质在物理热价和生物热价见表演7-1。
呼吸商 机体依靠呼吸功能从外界摄取氧,以供各种营养物质氧化分解的需要,同时也将代谢终生物CO2呼出体外,一定时间内机体的CO2产量与耗氧量的比值称为呼吸商(respiratory quotient, RQ)。各种营养物质在细胞内氧化供能属于细胞呼吸过程,因而双将各种营养物质氧化时的CO2产量与耗氧量的比值称为某物质的呼吸商。严格说来,应该以CO2和O2的克分子(mol)比值来表示呼吸商。但是,因为在同一温度和气压条件下,容积相等的不同气体,其分子数都是相等的,所以通常都用容积数(ml或L)来计算CO2与O2的比值,即:
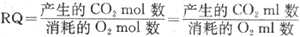
糖、脂肪和蛋白质氧化时,它们的CO2产量与耗氧量各不相同,三者的呼吸商也不一样。
因为各种营养物质无论在体内或体外氧化,它们的耗氧量与CO2产量都取决于各该物质的化学组成,所以,在理论上任何一种营养物质的呼吸商都可以根据它的氧化成终产物(CO2和H20)化学反应式计算出来的。
糖的一般分子式为(CH20)n,氧化时消耗的02和产生的C02分子数相等,呼吸商应该等于1。如上述葡萄糖氧化的反应式所示,C02产量与耗氧量均为6mol故:
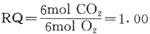
脂肪氧化时需要消耗更多的氧。在脂肪本身的分子结构中,氧的含量远较碳和氢少。因此,另外提供的氧不仅要用氧化脂肪分子中的碳,还要用来氧化其中的氢。所以脂肪的呼吸商将小于1。现以甘油三酸酯(triolein)为例:
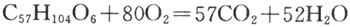
RQ=57molC02/80mol02=0.71
蛋白质的呼吸商较难测算,因为蛋白质在体内不能完全氧化,而且它氧化分解途径的细节,有些还不够清楚,所以只能通过蛋白质分子中的碳和氢被氧化时势需氧量和C02产量,间接算出蛋白质的呼吸商,其计算值为0.80。
在人的日常生活中,营养物质不是单纯的,而是糖、脂肪和蛋白质混合而成的(混合膳食)。所以,呼吸商常变动于0.71-1.00之间。人体在特定时间内的呼吸产要看哪种营养物质是当时的主要能量来源而定。若能源主要是糖类,则呼吸商接近于1.00;若主要是脂肪,则呼吸商接近于0.71。在长期病理性饥饿情况下,能源主要来自机体本身的蛋白质和脂肪,则呼吸商接近于0.80。一般情况下,摄取混合食物时,呼吸商常在0.85左右。
现将糖、脂肪和蛋白质三者的热价、氧热窬及呼吸商等数据列于表7-1,以供理解和测算能量代谢率之用。
表7-1 三种营养物质氧化时的几种数据
| 营养物质 |
产热量(kJ/g) |
耗氧量 |
CO2产量 |
氧热价 |
呼吸商 |
| 物理热价(用弹式热量计测得) |
生物热价(体内生物氧化什) |
营养学热价* |
|
| (L/g) |
(L/g) |
(kJ/J) |
(RQ) |
|
| 糖 |
17 |
17 |
16.7 |
0.83 |
0.83 |
21 |
1.00 |
| 蛋白质 |
23.5 |
18 |
16.7 |
0.95 |
0.76 |
18.8 |
0.80 |
| 脂肪 |
39.8 |
39.8 |
37.7 |
2.03 |
1.43 |
19.7 |
0.71 |
*营养学通常采用概数来计算食物的热价
影响呼吸商的其它因素:
机体的组织、细胞 不仅能同时氧化分解各种营养物质,而且也使一种营养物质转变为另一种营养物质。糖的转化为脂肪时,呼吸商可能变大,甚至超过1.00。这是由于当一部分糖转化为脂肪时,原来糖分子中的氧即有剩余,这些氧可能参加机体代谢过程中氧化反应,相应地减少了从外界摄取的氧量,因而呼吸商变大。反过来,如果脂肪转化为糖,呼吸商也可能低于0.71。这是由于脂肪分子中含氧比例小,当转化为糖时,需要更多的氧进入分子结构,因而机体摄取并消耗外界氧的量增多,结果呼吸商变小。另外,还有其它一些代谢反应也能影响呼吸商。例如,肌肉剧烈运动时,由于氧供不应求,糖酵解增多,将有大量乳酸进入血液。乳酸和碳酸盐作用的结果,会有大量由肺肺排出,此时呼吸商将变大。又如,肺过度通气、酸中毒等情况下,机体中与生物氧化无关的CO2大量排出,也可现呼吸大于1.00的情况。相反,肺通气不足、碱中毒等情况下,呼吸商将降低。
前已述,应该测出在一定时间内机体中糖、脂肪和蛋白质三者氧化分解的比例。为此。首先必须查清氧化了多少蛋白质,并且将氧化这些蛋白质所消耗的氧量和所产生的CO2从机体在该时间内的总耗氧量和总CO2产量中减去,算出糖和脂肪氧化(非蛋白质代谢)的CO2产量和耗氧量的比值,即非蛋白呼吸商(non-proteinrespiratory quotient,NPRQ),然后才有可能进一步查清糖和脂肪各氧化了多少克。
尿氮测定 尿中的氮物质主要是蛋白质的分解产物。因此可以通过尿氮来估算体内被氧化的蛋白质的数量。蛋白质的平均重量组成是:C50%,O223%,N16%,S1%。蛋白质中16%的N是完全随尿排出的。所以,1g 尿氮相当于氧分解6.25g蛋白质,测得的尿氮重量(g)乘以6.25,便相当于体内氧分解的蛋白质量。
非蛋白呼吸商 它是估算非蛋白代谢中糖和脂肪氧化的相对数量的依据。研究工作者早已按从0.707到1.00范围内的非蛋白呼吸产,算出糖和脂肪两者氧化的各自百分比以及氧热价(表7-2)。
表7-2 非蛋白呼吸商和氧热价
| 非蛋白呼吸商 |
氧 化 的 % |
氧热价(kJ/L) |
| 糖 |
脂肪 |
|
| 0.707 |
0.00 |
100.0 |
19.62 |
| 0.71 |
1.10 |
98.9 |
19.64 |
| 0.72 |
4.75 |
95.2 |
19.69 |
| 0.73 |
8.40 |
91.6 |
19.74 |
| 0.74 |
12.0 |
88.0 |
19.79 |
| 0.75 |
15.6 |
84.4 |
19.84 |
| 0.76 |
19.2 |
80.8 |
19.89 |
| 0.77 |
22.8 |
77.2 |
19.95 |
| 0.78 |
26.3 |
73.7 |
19.99 |
| 0.79 |
29.0 |
70.1 |
20.05 |
| 0.80 |
33.4 |
66.6 |
20.10 |
| 0.81 |
36.9 |
63.1 |
20.15 |
| 0.82 |
40.3 |
59.7 |
20.20 |
| 0.83 |
43.8 |
56.2 |
20.26 |
| 0.84 |
47,2 |
52.8 |
20.31 |
| 0.85 |
50.7 |
49.3 |
20.36 |
| 0.86 |
54.1 |
45.9 |
20.41 |
| 0.87 |
57.5 |
42.5 |
20.46 |
| 0.88 |
60.8 |
39.2 |
20.51 |
| 0.89 |
64.2 |
35.8 |
20.56 |
| 0.90 |
67.5 |
32.5 |
20.61 |
| 0.91 |
70.8 |
29.2 |
20.67 |
| 0.92 |
74.1 |
25.9 |
20.71 |
| 0.93 |
77.4 |
22.6 |
20.77 |
| 0.94 |
80.7 |
19.3 |
20.82 |
| 0.95 |
84.0 |
16.0 |
20.87 |
| 0.96 |
87.2 |
12.8 |
20.93 |
| 0.97 |
90.4 |
9.58 |
20.98 |
| 0.98 |
93.6 |
6.37 |
21.03 |
| 0.99 |
96.8 |
3.18 |
21.08 |
| 1.00 |
100.0 |
0.0 |
21.13 |
Lusk修订
间接测热法计算原则 实验测得的机体24小时内的耗氧量和CO2产量以及尿氮量,根据表7-1和7-2中相应的一些数据计算。首先,由尿氮量算出被氧分解的蛋白质量。由被氧化的蛋白质量从表7-1中算出其产热量、耗氧量和CO2产量;其次从总耗氧量和总CO2产量中减去蛋白质耗氧量和CO2产量,计算出非蛋白呼吸商。根据非蛋白呼吸商查表7-2的相应的非蛋白呼吸商的氧热价,计算出非蛋白代谢的产热量;最后,24小时产热量为蛋白质代谢的产热量与非蛋白代谢的产热量之和。此外,从非蛋白呼吸还可推算出参加代谢的糖和脂肪的比例。
间接测热法的计算方法举例
首先测定受试者一定时间内的耗氧量和CO2产量,假定受试者24小时的耗氧量为400L,CO2产量为340L(已换算成标准状态的气体容积)。另经测定尿氮排出量为12g。根据这些数据和查表7-1、7-2,计算24小时产热量,其步骤如下:(1)蛋白质氧化量=12×6.25=7g
产热量=18×75=1350kJ
耗氧量=0.95×75=71.25L
CO2产量=0.76×75=57L
(2)非蛋白呼吸商
非蛋白代谢耗氧量=400-71.25=328.75L
非蛋白代谢CO2产量=340-57=283L
非蛋白呼吸商=283/328.75=0.86
(3)根据非蛋白呼吸商的氧热价计算非蛋白代谢的热量
查表7-2,非蛋白呼吸商为0.86时,氧热价为20.41。所以,非蛋白代谢产热量=328.75×20.41=6709.8kj 。
(4)计算24小时产热量
24小时产热量=1350+6709.8=8059.8kJ
(蛋白质代(非蛋白代谢产热量)谢产热量)
计算的最后数值8059.8kJ就是该受试者24小时内的能量代谢率
耗氧量与CO2产量的测定方法及临床应用测定耗氧量和CO2产量的方法有两种:闭合式测定法和开放式测定法。
(1)闭合式测定法:在动物实验中,将受试动物置于一个密闭的能吸热的装置中。通过气泵,不断将定量的氧气送入装置。动物不断地摄取氧,可根据装置中氧量的减少计算出该动物在单位时间内的耗氧量。动物呼出的CO2则由装在气体回路中的CO2吸收剂吸收。然后根据实验前后CO2吸收剂的重量差,算出单位时间内的CO2产量。由耗氧量和CO2产量算出呼吸商。
临床上为了简便,通常只使用肺量计(图7-2)来测量耗氧量。该装置的气体中容器中装置氧气,受试者通过呼吸口瓣将氧气吸入呼吸器官。此时气体容器的上盖随吸气而下降,并由连于上盖的描笔记录在记录纸上。根据记录纸上的方格还可读出潮气量值。受试者的呼出气则通过吸收容器(呼出气中的CO2和水可除除掉)进入气体容器中,于是气体容器的上盖又复升高,描笔也了随之升高。由于受试者摄取了一定量的氧气,呼出气中CO2又被除掉,气体容器中的氧气量因而逐渐减少。描笔则记录出曲线逐渐下降的过程。在一定时间内(通常为6min),描笔的总下降高度,就是该时间内的耗氧量。
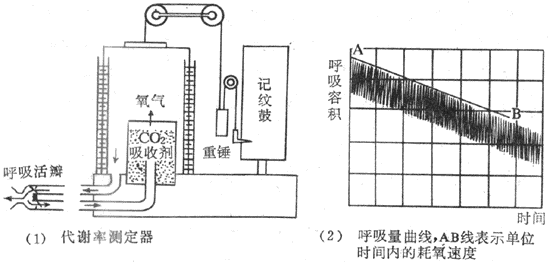
图7-2 肺量计结构模式图
(2)开放式测定法(气体分析法):它是在机体呼吸空气的条件下测定耗氧量和CO2产量的方法,所以称为开放法。其原理是,采集受试者一定时间内的呼出气,测定呼出气量并分析呼出气中氧和CO2的容积百分比。由于吸入气就是空气,所以其中氧和CO2的容积百分比不必另测。根据吸入气和呼出气中氧和CO2的容积百分比的差数,可算出该时间内的耗氧量和CO2排出量。
气体分析方法很多,最简便而又广泛应用的方法,是将受试者在一定时间内呼出气采集于气袋中,通过气量计测定呼气量,然后用气体分析器分析呼出气的组成成分,进而计算耗氧量和CO2产量,并算出呼吸商。
现举出一个气体分析实验例。
现某健康成人安静状态下的呼出气作气体分析,结果为:O2=16.26%;CO2=4.14%。呼出气量为1分钟5.2L(通常将呼出气量换算为不含水蒸气的标准状态值;也有换算为被水蒸气饱和的1个大气压、体温状态下的值)。空气的组成是:O2=20.96%;N=79.00%;CO2=0.04%。则:
受试者从每100ml通过肺的气体中吸收的氧气为:
20.96ml-16.26ml=4.7ml(或每升气体中的47ml)
(空气) (呼出气)
因为呼出气量为5.2L,则1分钟的耗氧量为:
47ml×5.2=244.4ml
100ml呼气中的CO2为:
4.14ml-0.04ml=4.1ml(或每升气体中的41ml)
(呼出气) (空气)
1分钟CO2排出量为:
41ml×5.2=213.2ml
RQ=213.1/244=0.85
所得呼吸商为受试者在安静状态下混合膳食代谢的呼吸商。
临床上和劳动卫生常采用简略法,即用气体分析法测得一定时间内的耗氧量和CO2产量,并求出呼吸商,并且不考虑蛋白质代谢部分,就根据非蛋白呼吸商表7-2查出呼吸商的氧热价,然后将氧热价乘以耗氧量,便得出该时间内的产热量。仍以上述间接测热法计算方法列举的例子为数据(见前),按此简略法来计算,则结果如下:
呼吸商=340/400=0.85
查表7-2,呼吸商0.85时的氧热价为20.36kJ,所以24小时的产热量=20.36×400=8144kJ。
这个数值与按完整的间接法计算得出的数值8059.8kJ是很近似的,误差都在1%-2%以下。而且在非蛋白呼吸商从0.70到1.00的范围内,氧热价也过变动于19.6-21.1之间。此法在实际工作中是可用的。
另一种更简便的简略法只利用肺量计测出受试者一定时间内(通常为6min)的耗氧量。受试者一般都吃混合膳食,所以通常将非蛋白呼吸商定为0.82,氧热价为20.20kJ。因此,测出一定时间内的耗氧量后,使可依下式来计算:
产热量=20.20×耗氧量(kJ)
二、影响能量代谢的因素
影响能量代谢的因素有肌肉活动、精神活动、食物的特殊动力作用和环境温度等。
(一)肌肉活动
肌肉活动对能量代谢的影响最为显着。机体任何轻微的活动都可提高代谢率。人在运动或劳动时耗量显着增加,因为肌肉活动需要补给能量,而能量则来自大量营养物质的氧化,导致机体耗氧量的增加。机体耗氧量的增加与肌肉活动的强度呈正比关系,耗氧量最多右达安静时的10-20倍。肌肉活动的强度称为肌肉工作的强度,也就是劳动强度。劳动强度通常用单位时间内机体的产热量来表示,也就是说,可以把能量代谢率作为评估劳动强度的指标。从表7-3可以看出劳动强度或运动时的能量代谢率的增长情况。
表7-3 运动或劳动时的能量代谢率
| 机体的状态 |
产 热 量 |
| 平均(kJ/m2·min) |
|
| 躺卧 |
2.73 |
| 开会 |
3.40 |
| 擦窗子 |
8.30 |
| 洗衣 |
9.98 |
■[此处缺少一些内容]■
由于随之出现的无意识的肌紧张以及刺激代谢的激素释放增多等原因,产热量可以显著增加。因此,在测定基础代谢率时,受试者必须摒除精神紧张的影响。
(三)食物的特殊动力作用
在安静状态下摄入食物后,人体释放的热量比摄入的食物本身氧化后所产生的热量要多。例如摄入能产100kJ热量的蛋白质后,人体实际产热量为130kJ,额外多产生了30kJ热量,表明进食蛋白质后,机体产热量超过了蛋白质氧化后产热量的30%。食物能使机体产生“额外”热量的现象称为食物的特殊动力作用(specific dynamic action)。糖类或脂肪的食物特殊动力作用为其产热量的4%-6%,即进食能产100kJ热量的糖类或脂肪后,机体产热量为104-106kJ。而混合食物可使产热量增加10%左右。这种额外增加的热量不能被利用来作功,只能用于维持体温。因此,为了补充体内额外的热量消耗,机体必须多进食一些食物补充这份多消耗的能量。
食物特殊动力作用的机制尚未完全了解。这种现象在进食后1h左右开始,并延续到7-8h。有人将氨基酸注入静脉内,可出现与经口给予相同的代谢率增值现象,这些事实使人们推想,食后的“额外”热量可能来源于肝处理蛋白质分解产物时“额外”消耗的能量。因此,有人认为肝在接脱氨基反应中消耗了能量可能是“额外”热量产生的原因。
(四)环境温度
人(裸体或只着薄衣)安静时的能量代谢,在20-30℃的环境中最为稳定。实验证明,当环境温度低于20℃时,代谢率开始有所增加,在10℃以下,代谢率便显着增加。环境温度低时代谢率增加,主要是由于寒冷刺激反射地引起寒战以及肌肉紧张增强所致。在20-30℃时代谢稳定,主要是由于肌肉松驰的结果。当环境温度为30-45℃时,代谢率又会逐渐增加。这可能是因为体内化学过程的反应速度有所增加的缘故,这时还有发汗功能旺盛及呼吸、循环功能增强等因素的作用。
三、基础代谢
基础代谢(basal metabolism)是指基础状态下的能量代谢。基础代谢率(basal metabolic rate,BMR)是指单位时间内的基础代谢,即在基础状态下,单位时间内的能量代谢。所谓基础状态是指人体处在清醒而又非常安静、不受肌肉活动、环境温度、食物及精神紧张等因素的影响时的状态。测定基础代谢率,要在清晨末进餐以前(即食后12-14h)进行。前一日晚餐最好是清淡菜肴,而且不要吃得太饱,这样,过了12-14h ,胃肠的消化和吸收活动已基本完毕,也排除了食物的特殊动力作用的影响。测定之前不应做剧烈的活动,而且必须静卧半小时以上。测定时平卧,全身肌肉要松驰,尽量排除肌肉活动的影响。这时还应要求受试者排除精神紧张的影响,如摒除焦虑、烦恼、恐惧等心理活动。室温要保持在20-25℃之间,以排除环境温度的影响。基本条件下的代谢率,比一般安静时的代谢率可低些(比清醒安静时低8%-10%)。基础代谢率以每小时,每平方米体表面积的产热量为单位,通常以kJ/m2·h来表示。要用每平方米体表面积而不用每公斤体重的产热量来表示,是因为基础代谢率的高低与体重并不成比例关系,而与体表面积基本上成正比。
表7-4 人体产热量和体表面积的关系
| 测定人数 |
平均体重(kg) |
体重范围 |
kJ/m2·24h |
| 6 |
48.7 |
40-50 |
3860 |
| 41 |
53.4 |
50-60 |
3827 |
| 164 |
64.5 |
60-70 |
3881 |
| 24 |
74.7 |
70-80 |
3868 |
| 8 |
83.7 |
80-90 |
3868 |
| 平均 |
|
|
3860 |
若以每公斤体重的产热量进行比较,则小动物每公斤体重的产热量要比大动物高得多。若以每平方米体表面积的产热量进行比较,则不论机体的大小,各种动物每平方米每24小时的产热量很相近。在人体上测定的结果也表明了这一事实(表7-4)。因此,用每平方米体表面积标准来衡量能量代谢是比较合适的。
受试者体表面积的测定繁琐而不易进行,鉴于体表面积与身高、体重之间有一定的相关关系,因此,有人对一定的人群作过测定后,从身高、体重推算出体表面计算的经验公式。最基本的是Meeh的算式:
S=KW2/3
式中,S为体表面积,W为体重(kg),K为不同种属动物的常数。
计算人的体表面积在DuBois的身长体重算式:
S=W0.425×H0.725×K
式中,S为体表面积(m2),W为体重(kg),H为身长。K为不同人种的常数。我国人的体表面积可根据下列Stevenson算式来计算:
体表面积(m2)=0.0061×身长(cm)+0.0128×体重(kg)-0.1529
另外,体表面积还可根据图7-3直接求出。其用法是,将受试者的身高和体重在相应两条列线的两点连成一直线,引直线与中间的体表面积列线的交点就是该人的体表面积。有意义的事实是:肺活量、心输出量、主动脉和气管的横截面、肾小球滤过率等都与体表面积有一定的比例关系。]
通常采用前述简略法来测定和计算基础代谢率。下面举一实例供参考。
某受试者,男性,20岁。在基础状态下,1h的耗氧量为15L。非蛋白呼吸商定为0.82,氧热价为20.20,所以1h的产热量为:
20.20×15=303kJ
此人的体表面积为1.5m2,从而1h内每平方米体表面积拉热量,即基本代谢率为:303÷1.5=201.8kJ/m2·h
20岁男子的正常基础代谢率为157.8kJ/m2·h,所以此人超出正常值的数字为(201.8-157.8)=44.0,超出正常值的百分数为:
44.0×100/157.8%=27.8%
即+27.8%

图7-3 体表面积测算用图
实际测定结果表明,基础代谢率随性别、年龄等不同而有生理变化。当甚情况相同时,男子的基础代谢率平均比女子的高;幼年人比成年人的高;年龄越大,代谢率越低,但是,同一个体的基础代谢率,只在测定时的条件完全符合前述的要求,则有不同时日重复测定的结果基本上无差异。这就反映了正常人的基础代谢率是相当稳定的。
关于我国正常人基础代谢率的水平,男女各年龄组的平均值如表7-5所示。
一般来说,基础代谢率的实际数值现上述正常的平均值比较,相差±10%-15%之内,无论较高或较低,都不属病态。当相差之数超过20%时,才有可能是病理变化。在各种疾病中,甲状腺功能的改变总是伴有基础代谢率异常变化。甲状腺功能低下时,基础代谢率将比正常值低20%-40%;甲状腺功能亢进时的基础代谢率将比正常值高出25%-80%(图7-4)。因此,基础代谢率的测量是临床诊断甲状腺疾病的重要辅助方法。其它如肾上腺皮质和垂体的功能低下时,基础代谢率也要降低。
表7-5 我国人正常的基础代谢率平均值(kJ/m2·h)
年龄 11-15 16-17 18-19 20-30 31-40 41-50 51 以上
男性 195.5 193.4 166.2 157.8 158.7 154.1 149.1
女性 172.5 181.7 154.1 146.4 142.4 142.4 138.6
当人体发热时,基础代谢率将升高。一般说来,体温每升高1℃,基础代谢率可升高13%。其它如糖尿病、红细胞增多症、白血病以及伴有呼吸困难的心脏病等,也伴有基础代谢率升高。当机体处于病理性饥饿时,基础代谢率将降低。其他如阿狄森病、肾病综合症以及垂体肥胖症也常伴有基础代谢率降低。
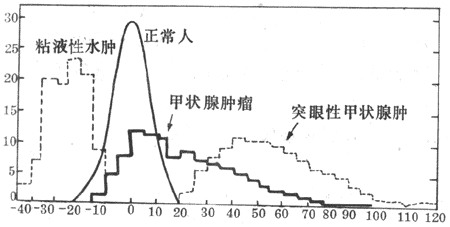
图7-4 甲状腺疾病患者的基础代谢与正常人基础代谢的比较
纵坐标表示受检人数;横坐标的0代表正常平均值,
-10表示比正常人平均值低10%,10表示高10%
第二节 体温及其调节
一、体温
人和高等动物机体都具有一定的温度,这就是体温。体温是机体进行新陈代谢和正常生命活动的必要条件。
(一)表层体温和深部体温
人体的外周组织即表层,包括皮肤、皮下组织和肌肉等的温度称为表层温度(shelltemperature)。表层温度不稳定,各部位之间的差异也不大。在环境温度为23℃时,人体表层最外层的皮肤温,如足皮肤温为27℃,手皮肤温为30℃。躯干为32℃,额部为33-34℃。四肢末稍皮肤温最低,越近躯干、头部,皮肤温越高。气温达32℃以上时,皮肤温的部位差将变小,在寒冷环境中,随着气温下降,手、足的皮肤温降低最显着,但头部皮肤温度变动相对较小。
皮肤与局部血流量有密切关系。凡是能影响皮肤血管舒缩的因素(如环境温度变化或精神紧张等)都能改变皮肤的温度。在寒冷环境中,由于皮肤血管收缩,皮肤血流量减少,皮肤温随之降低,体热散失因此减少。相反,在炎热环境中,皮肤血管舒张,皮肤血流量增加,皮肤温因而上升,同时起到了增强发散体热的作用。人情绪激动时,由于血管紧张度增加,皮肤温、特别是手的皮肤温便显著降低。例如手指的皮肤温可从30℃骤降到24℃。当然情绪激动的原因解除后,皮肤温会逐渐恢复。此外,当发汗时由于蒸发散热,皮肤温也会出现波动。
机体深部(心、肺、脑和腹腔内脏等处)的温度称为深部温度(coretemperature)。深部温度比表层温度高,且比较稳定,各部位之间的差异也较小。这里所说的表层与深部,不是指严格的解剖学结构,而是生理功能上所作的体温分布区域。在不同环境中,深部温度和表层温度的分布会发生相对改变。在较寒冷的环境中,深部温度分布区域较缩小,主要集中在头部与胸腹内脏,而且表层与深部之间存在明显的温度梯度。在炎热环境中,深部温度可扩展到四肢(图7-5)。
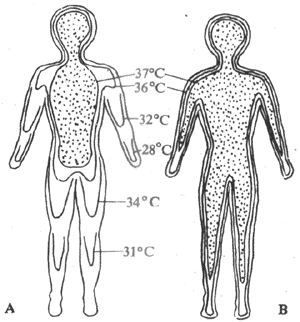
图7-5 在不同环境温度下人体体温分布图
A: 环境温度20℃ B:环境温度35℃
体温是指机体深部的平均温度。由于体内各器官的代谢水平不同,它们的温度略有差别,但不超过1℃。在安静时,肝代谢最活跃,温度最高;其次,是心脏和消化腺。在运动时则骨骼肌的温度最高。循环血液是体内传递热量的重要途径。由于血液不断循环,深部各个器官的温度会经常趋于一致。因此,血液的温度可以代表重要器官温度的平均值。
临床上通常用口腔温度、直肠温度和腋窝温度来代表体温。直肠温度的正常值为36.9-37.9℃,但易受下肢温度影响。当下肢冰冷时,由于下肢血液回流至髂静脉时的血液温度较低,会降低直肠温度;口腔温度(舌下部)平均比口腔温度低0.3℃,但它易受经口呼吸、进食和喝水等影响;腋窝温度平均比口腔温度低0.4℃.但由于腋窝不是密闭体腔,易受环境温度、出汗和测量姿势的影响,不易正确测定。
此外,食管温度比直肠温度约低 0.3℃。食管中央部分的温度与右心的温度大致相等,而且体温调节反应的时间过程与食管温度变化过程一致。所以,在实验研究中,食管温度可以作为深部温度的一个指标。鼓膜温度的变动大致与下丘脑温度的变化成正比,所以在体温调节生理实验中常常用鼓膜温度作为脑组织温度的指标。
(二)体温的正常变动
在一昼夜之中,人体体温呈周期性波动。清晨2-6时体温最低,午后1-6时最高。波动的幅值一般不超过1℃。体温的这种昼夜周期性波动称为昼夜节律或日周期(circadian rhythm)。
女子的基础体温随月经周期而发生变动。在排卵后体温升高,这咱体温升高一直持续至下次月经开始(图7-6)。这种现象很可能同性激素的分泌有关。实验证明,这种变动性同血中孕激素及其代谢产物的变化相吻合。
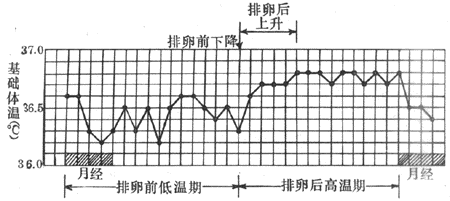
图7-6 女子的基础体温曲线
体温也与年龄有关。一般说来,儿童的体温较高,新生儿和老年人的体温较低。新生儿,特别是早产儿,由于体温调节机制发育还不完善,调节体温的能力差,所以他们的体温容易受环境温度的影响而变动。因此对新生儿应加强护理。
肌肉活动时代谢加强,产热量因而增加,结果可导致体温升高。所以,临床上应让病人安静一段时间以后再测体温。测定小儿体温时应防止哭闹。
此外,情绪激动、精神紧张、进食等情况对体温都会有影响;环境温度的变化对体温也有影响;在测定体温时,就考虑到这些情况。
二、体热平衡
如第一节所述,机体内营养物质代谢释放出来的化学能,其中50%以上以热能的形式用于维持体温,其余不足50%的化学能则载荷于ATP,经过能量转化与利用,最终也变成热能,并与维持体温的热量一起,由循环血液传导到机体表层并散发于体外。因此,机体在体温调节机制的调控下,使产热过程和散热过程处于平衡,即体热平衡,维持正常的体温。如果机体的产热量大于散热量,体温就会升高;散热量大于产热量则体温就会下降,直到产热量与散热量重新取得平衡时才会使体温稳定在新的水平。
(一)产热过程
机体的总产热量主要包括基础代谢,食物特殊动力作用和肌肉活动所产生的热量。基础代谢是机体产热的基础。基础代谢高产热量多;基础代谢低,产热量少。正常成年男子的基础代谢率约为170kJ/m2·h。成年女子约155kJ/m2·h在安静状态下,机体产热量一般比基础代谢率增高25%,这是由于维持姿势时肌肉收缩所造成的。食物特殊动力作用可使机体进食后额外产生热量。骨骼肌的产热量则变化很大,在安静时产热量很小。运动时则产热量很大;轻度运动如平行时,其产热量可比安静时增加3-5倍,剧烈运动时,可增加10-20倍。
人在寒冷环境中主要依靠寒战来增加产热量。寒战是骨骼肌发生不随意的节律性收缩的表现,其节律为9-11次/分。发生寒战的肌肉在肌电图上表现出一簇一簇的高波幅群放电,这是不同肌纤维的动作电位同步化的结果。寒战的特点是屈肌和伸肌同时收缩,所以基本上不做功,但产热量很高,发生寒战时,代谢率可增加4-5倍。机体受寒冷刺激时,通常在发生寒战之前,首先出现温度刺激性肌紧张(thermalmuscle tone)或称寒战前肌紧张(pre-shivering tone),此时代谢率就有所增加。以后由于寒冷刺激的持续作用,便在温度刺激性肌紧张的基础上出现肌肉寒战,产热量大大增加,这样就维持了在寒冷环境中的体热平衡。内分泌激素也可影响产热,肾上腺素和去甲肾上腺素可使产热量迅速增加,但维持时间短;甲状腺激素则使产热缓慢增加,但维持时间长。机体在寒冷环境中度过几周后,甲状腺激素分泌可增加2倍能上能下,代谢率可增加20-30%。
(二)散热过程
人体的主要散热部位是皮肤。当环境温度低于体温时,大部分的体热通过皮肤的辐射、传导和对流散热。一部分热量通过皮肤汗液蒸发来散发,呼吸、排尿和排粪也可散失一小部分热量(表7-6)。
表7-6 在环境温度为21℃时人体散热方式及其所占比例
| 散热方式 |
百 分 数 (%) |
| 辐射、传导、对流 |
70 |
| 皮肤水分蒸发 |
27 |
| 呼吸 |
2 |
| 尿、粪 |
1 |
1.辐射、传导和对流散热
辐射(radiation)散热:这是机体以热射线的形式将热量传给外界较冷物质一种散热形式。以此种方式散发的热量,在机体安静状态下所占比例较大(约占部散热量的60%左右)。辐射散热量同皮肤与环境间的温度差以及机体有效辐射面积等因素有关。皮肤温稍有变动,辐射散热量就会有很大变化。四肢表面积比较大,因此在辐射散热中有重要作用。气温与皮肤的温差越大,或是机体有效辐射面积越大,辐射的散热量就越多。
传导(conduction)和对流(convection)散热:传导散热是机体的热量直接传给同它接触的较冷物体的一种散热方式。机体深部的热量以传导方式传到机体表面的皮肤,再由后者直接传给同它相接触的物体,如床或衣服等。但由于此等物质是热的不良导体,所以体热因传导而散失的量不大。另外,人体脂肪的导热度也低,肥胖者皮下脂肪较多,女子一般皮下脂肪也较多,所以,他们由深部向表层传导的散热量要少些。皮肤涂油脂类物质,也可以起减少散热的作用。水的导热度较大,根据这个道理可利用冰囊、冰帽给高热病人降温。
对流散热是指通过气体或液体或交换热量的一种方式。人体周围总是绕有一薄层同皮肤接触的空气,人体的热量传给这一层空气,由于空气不断流动(对流),便将体热发散到空间。对流是传导散热的一种特殊形式。通过对流所散失的热量的多少,受风速影响极大。风速越大,对流散热量也越多,相反,风速越小,对流散热量也越少。
辐射、传导和对流散失的热量取决于皮肤和环境之间的温度差,温度差越大,散热量越多,温度差越小,散热量越少。皮肤温度为皮肤血流量所控制。皮肤血液循环的特点是,分布到皮肤的动脉穿透隔热组织(脂肪组织等),在乳头下层形成动脉网;皮下的毛细血管异常弯曲,进而形成丰富的静脉丛;皮下还有大量的动-静脉吻合支,这些结构特点决定了皮肤的血流量可以在很大范围内变动。机体的体温调节机制通过交感神经系统控制着皮肤血管的口径。增减皮肤血流量以改变皮肤温度,从而使散热量符合于当时条件下体热平衡的要求。
在炎热环境中,交感神经紧张度降低,皮肤小动脉命张,动-静脉吻合支开放,皮肤血流量因而大大增加(据测算,全部皮肤血流量最多可达到心输出量的12%)。于是较多的体热从机体深部被带到体表层,提高了皮肤温,增强了散热作用。
在寒冷环境中,交感神经紧张度增强,皮肤血管收缩,皮肤血流量剧减,散热量也因而大大减少。此时机体表层宛如一个隔热器,起到了防止体热散失的作用。此外,四肢深部的静脉是和动脉相伴走行的。这样的解剖结构相当于一个热量逆流交换系统。深部静脉呈网状围绕着动脉。静脉血温较低,而动脉血温度较高。两者之间由于温度差而进行热量交换。逆流交换的结果,动脉血带到末稍的热量,有一部分已被静脉血带回机体深部。这样就减少了热量的散失。如果机体处于炎热环境中,从皮肤返回心脏的血液主要由皮肤表层静脉来输送,此时逆流交换机制将不再起作用(图7-7)。
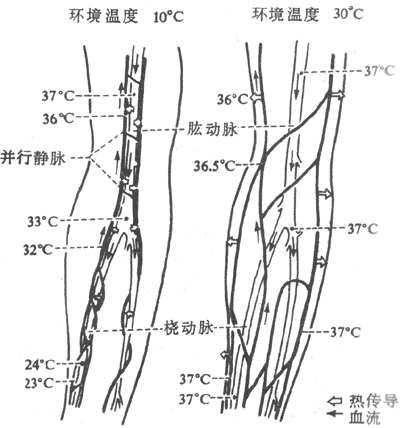
图7-7 上肢的逆流热量交换
环境温度降低时,热量由肱动脉传向它周围的静脉,
动脉血温度因此下降到24℃。环境温度升高时,热量由表层静脉发散
衣服覆盖的皮肤表层,不易实现对流,棉毛纤维间的空气不易流动,这类情况都有利于保温。增加衣着以御寒,就是这个道理。
2.蒸发散热 在人的体温条件下,蒸发(evaporation)1g水分可使机体散失2.4kJ热量。当环境温度为21℃时,大部分的体热(70%)靠辐射、传导和对流的方式散热,少部分的体热(29%)则由蒸发散热;当环境温度升高时,皮肤和环境之间的温度差变小,辐射、传导和对流的散热量减小,而蒸发的散热作用则增强;当环境温度等于或高于皮肤温度时,辐射、传导和对流的散热方式就不起作用,此时蒸发就成为机体唯一的散热方式。
人体蒸发有二种形式:即不感蒸发(insesibleperspiration)和发汗(sweating)。人体即使处在低温中,没有汁液分泌时,皮肤和呼吸道都不断有水分渗也而被蒸发掉,这种水分蒸发称为不感蒸发,其中皮肤的水分蒸发又称为不显汗,即这种水分蒸发不为人们所觉察,并与汁腺的活动开关。在室温30℃以下时,不感蒸发的水分相当恒定,有12-15g/h·m2水分被蒸发掉,其中一半是呼吸道蒸发的水分;另一半的水分是由皮肤的组织间隙直接渗出而蒸发的。人体24h的不感蒸发量为400-600ml。婴幼儿的不感蒸发的速率比成从大,因此,在缺水时婴幼儿更容易造成严重脱水。不感蒸发是一种很有成效的散热途径,有些动物如狗,虽有汁腺结构,但在高温环境下也不能分泌汁液,此时,它必须通过热喘呼吸(panting)由呼吸道来增强蒸发散热。
发汗汗腺分泌汁液的活动称为发汗。发汗是可以意识到的有明显的汗液分泌,因此,汁液的蒸发又称为可感蒸发。
人在安静状态下,当环境温度达30℃左右时便开始发汗。如果空气湿度大,而且着衣较多时,气温达25℃便可引起人体发汗。人进行劳动或运动时,气温虽在20℃以下,亦可出现发汗,而且汗量往往较多的。
汗液中水分占99%,而固体成分则不到1%,在固体成分中,大部分为氯化钠,也有少量氯化钾、尿素等。同血浆相比,汗液的特点是:氯化钠的浓度一般低于血浆;在高温作业等大量出汗的人,汗液中可丧失较多的氯化钠,因此应注意补充氯化钠。汗液中葡萄糖的浓度几乎是零;乳酸浓度主于血浆;蛋白质的浓度为零。实验测得在汗腺分泌时,分泌管腔内的压力高达37.3kPa(250mmHg)以上。这表明汗液不是简单的血浆滤出液,而是由汗腺细胞主动分泌的。大量的乳酸是腺细胞进入分泌活动的产物。刚刚从汗腺细胞分泌出来的汗液,与血浆是等渗的,但在流经汗腺管腔时,由于钠和氯被重吸收,所以,最后排出的汗液是低渗的。汗液中排出的钠量也受醛固醇的调节。下因为汗液是低渗的,所以当机体因大量发汗而造成脱水时,可导致高渗性脱水。
发汗是反射活动。人体汗腺接受交感胆碱能纤维支配,所以乙酰胆碱对小汗腺有促进分泌作用。发汗中枢分布在从脊髓到大脑皮层的中枢神经系统中。在正常情况下,起主要作用是是下丘脑的发汗中枢,它很可能位于体温调节中枢之中或其附近。
在温热环境下引起全身各部位的小汗腺分泌汗液称为温热性发汗。始动温热性发汗的主要因素有:①温热环境刺激皮肤中的瘟觉感受器,冲动传入至发汗中枢,反射性引起发汗;②温热环境使皮肤血液被加温,被加温的血液流至下丘脑发汗中枢的热敏神经元,可引起发汗。温热性发汗的生理意义在于散热。若每小时蒸发1.7L汗液,就可使体热散发约4200kJ的热量。但是,如果汗水从身上滚落或被擦掉而未被蒸发,则无蒸发散热作用。
发汗速度受环境温度和湿度影响。环境温度越高,发汗速度越快。如果在高温环境中时间太长,发汗速度会因汗腺疲劳而明显减慢。湿度大,汗液不易被蒸发,体热因而不易蒸发,体热因而不易散失。此外,风速大时,汗液易蒸发,汗液蒸发快,容易散热而使发汗速度变小。
劳动强度也影响发汗速度。劳动强度大,产热量越多,发汗量越多。
精神紧张或情绪激动而引起地发汗称为精神性发汗。主要见于掌心、脚底和腋窝。精神性发汗的中枢神经可能在大脑皮层运动区。精神性发汗在体温调节中的作用不大。
三、体温调节
恒温动物包括人,有完善的体温调节机制。在外界环境温度改变时,通过调节产热过程和散热过程,维持体温相对稳定。例如,在寒冷环境下,机体增中产热和减少散热;在炎热环境下,机体减少产热和增加散热,从而使体温保持相对稳定。这是复杂的调节过程,涉及感受温度变化的温度感觉器,通过有关传导通路把温度信息传达到体温调节中枢,经过中枢整合后,通过自主神经系统调节皮肤血流量、竖毛肌和汗腺活动等;通过躯体神经调节骨骼肌的活动,如寒战等;通过内分泌系统,改变机体的代谢率。
体温调节是生物自动控制系统的实例。如图7-8所示,下丘脑体温调节中枢,包括调定点(set point)神经元在内,属于控制系统。它的传出信息控制着产热器官如肝、骨骼肌以及散热器官如皮肤血管、汗腺等受控系统的活动,使受控对象——机体深部温度维持一个稳定水平。而输出变量体温总是会受到内、旬环境因素干扰的(譬如机体的运动或外环境气候因素的变化,如气温、湿度、风速等)。此时则通过温度检测器——皮肤及深部温度感受器(包括中枢温度感受器)将干扰信息反馈于调定点,经过体温调节中枢的整合,再调整受控系统的活动,仍可建立起当时条件下的体热平衡,收到稳定体温的效果。
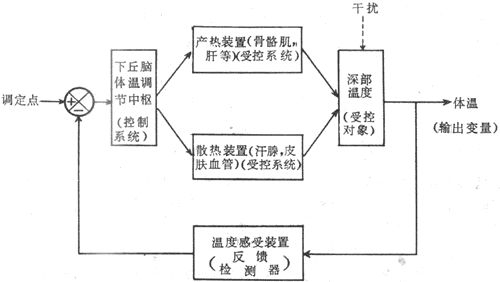
图7-8 体温调节自动控制示意图
(一)温度感受器
对温度敏感的感受器称为温度感受器,温度感受器分为外周温度感受器和中枢温度感受器。
外周温度感受器在人体皮肤、粘膜和内脏中,温度感受器分为冷感受器和温觉感受器,它们都是游离神经末稍的。当皮肤温度升高时,温觉感受器兴奋,而当皮肤温度下降时,则冷感受器兴奋。从记录温度感受器发放冲动可看到,温觉感受器和冷觉感受器在28℃时发放冲动频率最高,而温觉感受器则在43℃时发放冲动频率最高。当皮肤温度偏离这两个温度时,两种感受器发放冲动的频率都逐渐下降(图7-9)。此外,温度感受器对皮肤温度变化速率更敏感。
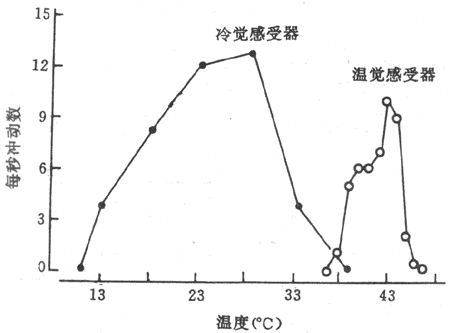
图7-9 大鼠阴囊皮肤冷觉和温觉感受器
内脏器官也有温度感受器。有人将电热器埋藏在差点羊腹腔内 并加温至43-44℃,观察到羊的呼吸频率和蒸发散热迅速增加,加热3-5分钟后,动物开始喘息,使下丘脑温度下降。说明内脏温度升高可引起明显的散热反应。
中枢温度感受器在脊髓、延髓、脑干网状结构及下丘脑中有温度感受器。
用改变脑组织温度的装置(变温管),对不麻醉或麻醉的兔、猫或狗等的下丘脑前部进行加温或冷却,发现在视前区-下丘脑前部(preoptic anterior hypothalamus,PO/AH)加温,可引起动物出现喘息和出汗等散热反应,而局部冷却则引起产热量增加,说明PO/AH本身就可调节散热和产热这两种相反的过程。用电生理方法记录PO/AH中存在着热敏神经元(warm-sensitive neuron)和冷敏神经元(cold-sensitiveneuron)。前者的放电频率随局部温度的升高而增加,而后者的放电频率则随着脑组织的降温而增加(图7-10,7-11)。实验证明,局部脑组织温度变动0.1C,这两种温度敏感神经元的放电频率就会反映出来,而且不出现适应现象。
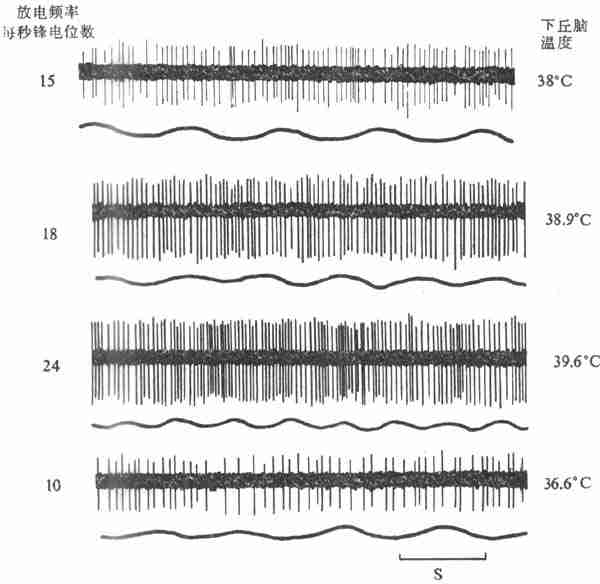
图7-10下丘脑局部加温时热敏神经元放电的记录(上)和呼吸曲线(下)(猫)
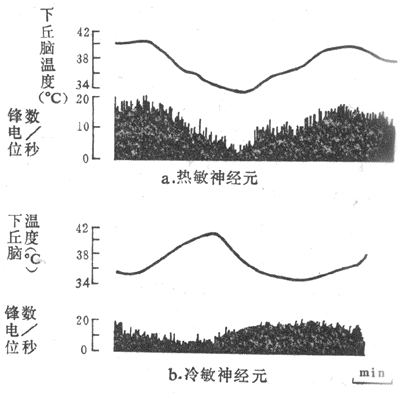
图7-11由下丘脑视前区导出的温度敏感神经元的放电活动
脊髓中也有温度敏感神经元。冷却轻度麻醉狗的颈、胸髓或胸腰髓,则动物出现皮肤血管收缩和寒战等体温调节反应。这时,切断被冷却部位的后根或高位切断脊髓,血管反应和寒战也不消失。加温脊髓,则引起皮肤血管舒张和热喘呼吸,寒战受到抑制。另外,据谓脊髓中传导温度信息的上行性神经元的纤维前侧侧索中走行,它将信息发送给PO/AH。
延髓中也存在着温度敏感神经元。皮肤、脊髓及中脑的传入温度信息都会聚于延髓温度敏感神经元;而延髓也接受来自PO/AH的信息,并且向PO/AH输送信息。
脑干网状结构也有对局部温度变化发生反应的神经元,它接受发生皮肤、脊髓的温度信息,并且向PO/AH输送温度信息。
(二)体温调节中枢
根据多种恒温动物脑的分段切除实验看到,切除大脑皮层及部分皮层下结构后,只要保持下丘脑及其以下的神经结构完整,动物虽然在行为方面可能出现一些欠缺,但仍具有维持恒定体温的能力。如进一步破坏下丘脑,则动物不再具有维持体温相对恒定的能力。这些事实说明,调节体温的基本中枢在下丘脑。下丘脑局部破坏或电刺激等实验观察到,PO/AH破坏,则散热反应消失,体温升高;刺激之,则引起散热反应,而且寒战受到抑制;而破坏下丘脑后部,体温下降,产热反庆受抑制;刺激之,则引起寒战。据此得出结论,下丘脑前部是散热中枢,而下丘脑后部是产热中枢,但是,这两种实验方法比较粗糙,因此得出来的结论也较精细的实验方法观察到的结果不相符。
前已述及, PO/AH就有热敏神经元和冷敏神经元,分别调节散热和产热反应。下丘脑以外的脑细胞也有类似的两种神经元存在。看来没有明确定位的产热中枢和散热中枢。体温调节是涉及多方输入温度信息和多系统的传出反应,因此是一种高级的中枢整合作用。视前区-下丘脑前部应是体温调节的基本部位。下丘脑前部的热敏神经元和冷敏神经元既能感受它们所在部位的温度变化,又能过传入的温度信息进行整合。因此,当外界环境温度改变时,可通过①皮肤的温、冷觉感受器的刺激,将温度变化的信息沿躯体传入神经经脊髓到达下丘脑的体温调节中枢;②外界温度改变可通过血液引起深部温度改变,并直接作用于下丘脑前部;③脊髓和下丘脑以外的中枢温度感受器也将温度信息传给下丘脑前部。通过下丘脑前部和中枢其它部位的整合作用,由下述三条途径发出指令调节体温:①通过交感神经系统调节皮肤血管舒缩反应和汗腺分泌;②通过躯体神经改变骨骼肌的活动,如在寒冷环境时的寒战等;③通过甲状腺和肾上腺髓质的激素分泌活动的改变来调节机体的代谢率。有人认为,皮肤温度感受器兴奋主要调节皮肤血管舒活动和血流量;而深部温度改变则主要调节发汗和骨骼肌的活动。通过上述的复杂调节过程,使机体在外界温度改变时能维持体温相对稳定。
调定点学说 此学说认为,体温的调节类似于恒温器的调节,PO/AH中有个调定点,即规定数值(如37℃)。如果偏离此规定数值,则由反馈系统将偏离信息输送到控制系统,然后经过对受控系统的调整来维持体温的恒定。通常认为,PO/AH中的温度敏感神经元可能在体温调节中起着调定点的作用。例如,此学说认为,由细菌所致的发热是由于热敏神经元的阈值因受到热原(pyrogen)的作用而升高,调定点上移(如39℃)的结果。因此,发热反应开始先出现恶寒战栗等产热反应,直到体温升高到39℃以上时才出现散热反应。只要致热因素不消除,产热与散热两个过程就继续在此新的体温水平上保持着平衡。应该指出的是,发热时体温调节功能并无阻碍,而只是由于调定点上移,体温才被调节到发热水平。
单胺物质对体温调节的作用在哺乳动物下丘脑的与体温调节有关的神经末稍中含有丰富的单胺物质。60年代初,用狗、猫、猴做的实验证明,用5-羧色胺灌注动物的脑室或微量注入于下丘脑,动物的体温上升,同时伴有血管收缩反应和寒战;而去甲肾上肾素则使动物的体温降低0.5-2℃,同时伴有外周血管舒张。根据这类实验,提出了体温调节的单胺学说,此学说认为,5-羧色胺和去甲肾上腺素这两种物质在量上的动态平衡可保持体温的恒定。但目前认为,这两种物质对体温调节中枢的活动只能起到调整的作用,而对于体温的恒定水平没有决定作用。
参考资料
1.徐丰彦,张镜如主编.人体生理学.北京;人民卫生出版社,1989
2.Patton HD,FuchsAE,Hille B,Scher RS,Textbook of Physiology 21th ed,WB SaundersCO,Philadelphia,1989
3.Ganong WF.Review of MedicalPhysiology 13th ed,Lange Medical Publications,LosAltos,Calfornia,1991
4.Emslie-Smith D,PatersonCR,Scratcherd T,Read NW.Textbook of Physiology,Scratchill Livingstone EdinburghLondon Melbourne and New York,1988
5.Mountcastle VB,MedicalPhysiology 14th ed,CV Mosby Co,St Louis 1980

 中医世家
中医世家 浦 标 网
浦 标 网 河南大学精品课程
河南大学精品课程 图书资料室
图书资料室