第一节 细胞因子的概述
一、胞因子的概念
机体的免疫细胞和非免疫细胞能合成和分泌小分子的多肽类因子,它们调节多种细胞生理功能,这些因子统称为细胞因子(cytokines)。细胞因子包括淋巴细胞产生的淋巴因子和单核巨噬细胞产生的单核因子等。目前已知白细胞介素(interleukin,IL),干扰素(interferon,IFN)、集落刺激因子(colony stimulating factor,CSF)、肿瘤坏死因子(tumornecrosis factor,TNF)、转化生长因子(transforming growth foctor,TGF-β)等均是免疫细胞产生的细胞因子,它们在免疫系统中起着非常重要的调控作用,在异常情况下也会导致病理反应。
研究细胞因子有助于阐明分子水平的免疫调节机制,有助于疾病的预防、诊断和治疗,特别是利用细胞因子治疗肿瘤、感染、造血功能障碍、自身免疫病等,已收到初步疗效,具有非常广阔的应用前景。
二、细胞因子的命名
(一)白细胞介素
在1979年第二届淋巴因子的国际会议上,将介导白细胞间相互作用的一些细胞因子命名为白细胞介素(IL),并以阿拉伯数字排列,如IL-1、IL-2、IL-3。随着分子免疫学的研究进展,不断有新的IL被命名,迄今已正式命名到IL-15,可以预期,还会有更多的IL被发现。目前的研究发现,许多IL不仅介导白细胞相互作用,还参与其它细胞的相互作用,如造血干细胞、血管内皮细胞、纤维母细胞、神经细胞、成骨和破骨细胞等的相互作用(表4-1)。
表4-1 白细胞介素的特性(IL)
| IL |
曾用名称 |
产生细胞 |
效应 |
| 1 |
淋巴细胞活化因子(LAF) |
单核巨噬细胞,树突状细胞,纤维母细胞内皮细胞 |
T和B细胞的增殖和分化,刺激造血细胞,参予炎症反应 |
| 2 |
T细胞生长因子 |
活化的T细胞 |
T和B细胞的增殖分化,增强NK细胞,单核细胞杀伤活性 |
| 3 |
多集落刺激因子(multi-CSF) |
活化的T细胞 |
多能造血干细胞增殖,促进肥大细胞,嗜酸,嗜碱性粒细胞增殖与分化 |
| 4 |
B细胞刺激因子(BSF-1) B细胞生长因子(BCGF-1) |
活化的T细胞 |
B和T细胞增殖,刺激造血祖细胞增殖与分化,诱导lgE、lgG产生 |
| 5 |
B细胞生长因子-Ⅱ (BCGF-Ⅱ) |
活化的T细胞 |
促进B细胞增殖与分化,促进嗜酸性粒细胞增殖与分化,诱导lgA产生 |
| 6 |
B细胞刺激因子-2(BSF-2) B细胞分化因子(BCDF) |
淋巴细胞 单核细胞 纤维母细胞 |
促进B细胞分化、促进肝细胞产生急性期蛋白,抑制乳腺癌细胞、刺激骨髓瘤细胞、刺激造血细胞,参与炎症 |
| 7 |
淋巴细胞生素(LPO) |
骨髓及胸腺基质细胞 |
促进前T、前B细胞增殖,促进成熟T细胞生长,促进血小板生成 |
| 8 |
中性粒细胞趋化因子(NCF) 粒细胞活化因子(NAF) |
单核巨噬细胞 血管内皮细胞 |
中性粒细胞活化和趋化作用,T细胞趋化作用,促进血管生成,参与炎症 |
| 9 |
P40 肥大细胞生长增强活性T 细胞生长因子(TCGFⅢ) |
活化的T 细胞 |
促进TH产生细胞因子,促进肥大细胞增殖,刺激造血细胞 |
| 10 |
细胞因子合成抑制因子(CSIF) |
活化的T细胞,B细胞单核巨噬细胞 |
抑制TH产生细胞因子,促朝进胸腺细胞增殖,促进B细胞增殖 |
| 11 |
(一) |
骨髓基质细胞 |
促进B细胞分化,刺激造血细胞,促进血小板生成 |
| 12 |
细胞毒性淋巴细胞成熟因子(CLMF) |
B细胞 |
促进TC,NK,LAK细胞杀伤功能,透导细胞免疫 |
| 13 |
P600 |
活化的T细胞 |
抑制细胞因子分泌和表达,刺激B细胞增殖和CD23表达,透导lgE产生 |
(二)集落刺激因子
在进行造血细胞的体外研究中,发现一些细胞因子可刺激不同的造血干细胞在半固体培养基中形成细胞集落,这类因子被命名为集落刺激因子(CSF)。根据它们的作用范围,分别命名为粒细胞CSF(G-CSF),巨噬细胞CSF(M-CSF),粒细胞和巨噬细胞CSF(GM-CSF)和多集落刺激因子(multi-CSF,又称IL-3)。不同发育阶段的造血干细胞起促增殖分化的作用,是血细胞发生必不可少的刺激因子。广义上,凡是刺激造血的细胞因子都可统称为CSF,例如刺激红细胞生成素(erythropoictin,Epo)、刺激造血干细胞的干细胞因子(stem cellfactor,SCF)、可刺激胚胎干细胞的白血病抑制因子(leukemia inhibitory factor,LIF)等均有集落刺激活性。此外,CSF也作用于多种成熟的细胞,促进其功能具有多相性的作用(表4-2)。
表4-2 集落刺激因子的特性
| 细胞因子 |
产生细胞 |
效应 |
| Multi-CSF |
活化的T细胞 |
刺激造血干细胞增殖,促进肥大细胞,嗜酸、嗜碱粒细胞增殖分化 |
| GM-CSF |
活化的T细胞,巨噬细胞,纤维母细胞等 |
刺激粒细胞,巨噬细胞集落形成刺激粒细胞功能 |
| G-CSF |
纤维母细胞,骨髓基质细胞,膀胱癌细胞株等 |
刺激粒细胞集落,刺激粒细胞功能 |
| M-CSF |
巨噬细胞 |
刺激巨噬细胞集落、刺激粒细胞功能,降低血胆固醇 |
| SCF |
纤维母细胞,骨髓和胸腺的基质细胞 |
刺激髓系、红系、巨核系及淋巴系造血祖细胞 |
| Epo |
肾细胞 |
刺激红系造血祖细胞 |
| LIF |
基质细胞、单核细胞 |
促进某些白血病细胞株的分化促进胚胎干(ES)细胞的增殖,抑制ES细胞的分化 |
(三)干扰素
干扰素(IFN)是最先发现的细胞因子,早在1957年,lssacs等人发现病毒感染的细胞产生一种因子,可抵抗病毒的感染,干扰病毒的复制,因而命名为干扰素。根据其来源和结构,可将IEN分为IFN-α、IFN-β、IFN-γ,它们分别由白细胞、纤维母细胞和活化T细胞产生。IFN-α为多基因产物,有十余种不同亚型,但它们的生物活性基本相同。IFN除有抗病毒作用外,还有抗肿瘤、免疫调节、控制细胞增殖及引起发热等作用。
(四)肿瘤坏死因子
TNF是一类能直接造成肿瘤细胞死亡的细胞因子,根据其来源和结构分为两种,即TNF-α和TNF-β.前者由单核巨噬细胞产生;后者由活化的T细胞产生,又名淋巴毒素(lymphotoxin)。TNF除有杀肿瘤细胞作用外,还可引起发热和炎症反应,大剂量TNF-α可引起恶液质,呈进行性消瘦,因而TNF-α又称恶液质素(cac hectin)。
(五)淋巴因子
由活化的淋巴细胞产生的细胞因子都可称为淋巴因子(lymphokine),如IL-2、3、4、5、6、7、8、9、10、11、12、13,TNF-β,IFN-γ等均为淋巴因子。
(六)单核因子
由单核已噬细胞产生的细胞因子统称单核因子(monokine),如IL-1、6、8,TNF-α、IFN-α等。
三、细胞因子的作用特点
目前发现并正式命名的细胞因子有数十种,每种细胞均有其独特的、起主要作用的生物学活性。尽管种类繁多、产生细胞和作用细胞多样、生物学活性广泛、发挥作用的机制不同,但众多的细胞因子具有以下共同的特性:
1.天然细胞因子是由细胞产生的 正常的静息或休止(resting)状态的细胞必须经过激活后才能合成和分泌细胞因子。通常是由抗原、丝裂原或其它刺激物激活免疫细胞和相关细胞,6~8小时后细胞培养上清中即可检测出细胞因子,于24~72小时期间细胞因子水平最高。但是有些细胞株不需外源刺激就可以自发地分泌某些细胞因子。
2.细胞因子的产生和作用具有多向性(pleiotropism)即单一刺激如抗原、丝裂原、病毒感染等可使同一种细胞分泌多种细胞因子,而一种细胞因子由多种不同类型的细胞产生可作用于多种不同类型的靶细胞。
3.细胞因子的合成和分泌过程是一种自我调控的过程通常情况下,细胞因子极少储存,即不以前体形式贮存在细胞内,而是经过适当刺激后迅速合成,一旦合面后便分泌至细胞外以发挥生物学作用,刺激消失后合成亦较快地停止并被迅速降解。
4.为低分子量的分泌型蛋白质常被糖基化。分子量大小不等,大多数为15~30kD,小者仅8~10kD,一般不超过80kD。
5.细胞因子需与靶细胞上的高亲和力受体特异结合后才发挥生物学效应。
6.生物学效应极强 细胞因子在pM(10-12M)水平就能发挥显著的生物学效应。这与细胞因子与靶细胞表面特异性受体之间亲和力极高有关,其解离常数在10-12~10-10M之间。
7.单一细胞因子可具有多种生物学活性,但多种细胞因子也常具有某些相同或相似的生物学活性。
8.主要参与免疫反应和炎症反应影响反应的强度和持续时间的长短。涉及到感染免疫、肿瘤免疫、自身免疫、移植免疫等诸多方面。
9.以非特异性方式发挥生物学作用且不受MHC限制。
10.某种细胞因子对靶细胞作用的强弱取决于细胞因子的局部浓度,靶细胞本身的类型(即作用于自身产生细胞)和旁分泌方式(paracrine,即作用于邻近的靶细胞)短暂性地产生并在局部发挥作用。
11.天然细胞因子大多是在近距离发挥局部作用 大多是通过自分泌方式(autocrine,即作用于自身产生细胞)和旁分泌方式(paracrine,即作用于邻近的靶细胞)短暂性地产生并在局部发挥作用。
12.细胞因子的作用并不是孤立存在的,它们之间通过合成分泌的相互调节,受体表达的相互调控、生物学效应的相互影响而组成细胞因子网络(addidveeffect)也可以取得协同效应(synergy),甚至取得两种细胞因子单用时所不具有的新的独特的效应。
第二节 细胞因子及其受体的结构
一、细胞因子的分子结构
不同细胞因子之间的结构上有很大的差异,一般,多数细胞因子为小分子多肽,分子量不超过60kD,多由100个左右的氨基酸组成。不同细胞因子之间无明显的氨基酸序列的同源性。
多数细胞因子以单体形式存在,少数因子如IL-5、IL-12、M-CSF、TGF-β等以双体形式存在。
给大多数细胞因子带有糖基,但这些糖基多与细胞因子的生物活性无关,可能起延长细胞因子体内半衰期的作用。
二、细胞因子受体
细胞因子都是通过与靶细胞表面高亲合力的特异性受体结合后才能发挥其生物学效应的。细胞因子受体与其它膜表面受体一样,均由3个功能区组成,即膜外区(细胞因子结合区)。跨膜区(疏水性氨基酸富有区)和膜内区(信号传导区)。细胞因子受体存在有单链、双链或三链不同形式的结构。最近的研究发现,有些细胞因子受体共同使用一条多肽链,如IL-3、IL-5和GM-CSF共同使用同一β链,IL-2、IL-4和IL-7共同使用同一γ链。由于细胞因子在受体水平存在相似性,因而会使用共同的信号传导途径,发挥类似的生物学效应。根据细胞因子受体膜外区的氨基酸序列,可将其主要分为三个受体家族:
(一)造血生长因子受体家族(HPR)
大部分细胞因子如IL-2、3、4、5、6、7、9等的受体均属于这一家族,其典型结构特点是含有Trp-Ser-X-Trp-Ser(W-S-X-W-S)的五联保守序列,与细胞因子结合功能密切相关。
(二)lg超家族
IL-1受体、M-CSF受体等属于这一家族,IL-6受体同时含有lg超家族和HPR家族两个结构区。这一超家族的特点是均在膜外区含有lg样的分子构型,每个lg样功能区由100个左右的氨基酸组成,通过二硫键形成稳定的发夹样反平行的β片层折叠结构。
(三)干扰素受体超家族
干扰素α和β共用同一个受体,与干扰素γ受体的结构有类似之外,均含有一段200个氨基酸的保守序列,其中4个半胱氨酸是共有的。
第三节 细胞因子的生物学活性
细胞因子具有非常广泛的生物学活性,包括促进靶细胞的增殖和分化,增强抗感染和细胞杀伤效应,促进或抑制其它细胞因子和膜表面分子的表达,促进炎症过程,影响细胞代谢等。
一、免疫细胞的调节剂
免疫细胞之间存在错综复杂的调节关系,细胞因子是传递这种调节信号必不可少的信息分子。例如在T-B细胞之间,T细胞产生IL-2、4、5、6、10、13,干扰素γ等细胞因子刺激B细胞的分化、增殖和抗体产生;而B细胞又可产生IL-12调节TH1细胞活性和TC细胞活性。在单核巨噬细胞与淋巴细胞之间,前者产生IL-1、6、8、10,干扰素α,TNF-α等细胞因子促进或抑制T、B、NK细胞功能;而淋巴细胞又产生IL-2、6、10,干扰素γ,GM-CSF,巨噬细胞移动抑制因子(MIF)等细胞因子调节单核巨噬细胞的功能。许多免疫细胞还可通过分泌细胞因子产生自身调节单核巨噬细胞的功能。许多免疫细胞还可通过分泌细胞因子产生自身调节作用。例如T细胞产生的IL-2可刺激T细胞的IL-2受体表达和进一步的IL-2分泌,TH1细胞通过产生干扰素γ抑TH2细胞的细胞因子产生。而TH2细胞又通过IL-10、IL-4和IL-13抑制TH1细胞的细胞因子产生。通过研究细胞因子的免疫网络调节,可以更好地理解完整的免疫系统调节机制,并且有助于指导细胞因子做为生物应答调节剂(biologicalresponsemodifier,BRM)应用于临床治疗免疫性疾病。
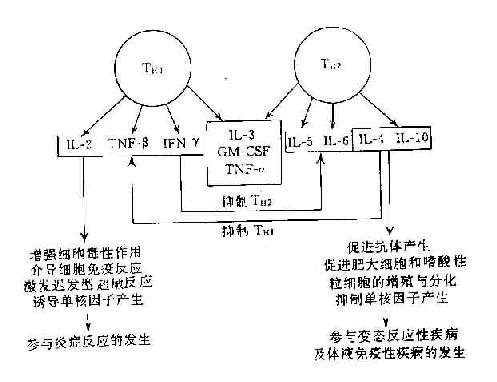
图4-1 细胞因子与TH1、TH2的相互关系
二、免疫效应分子
在免疫细胞针对抗原(特别是细胞性抗原)行使免疫效应功能时,细胞因子是其中重要效应分子之一。例如TNFα和TNFβ可直接造成肿瘤细胞的凋零(apoptosis),使瘤细胞DNA断裂,细胞萎缩死亡;干扰素α、β、γ可干扰各种病毒在细胞内的复制,从而防止病毒扩散;LIF可直接作用于某些髓性白血病细胞,使其分化为单核细胞,丧失恶性增殖特性。另有一些细胞因子通过激活效应细胞而发挥其功能,如IL-2和IL-12刺激NK细胞与TC细胞的杀肿瘤细胞活性。与抗体和补体等其它免疫效应分子相比,细胞因子的免疫效应功能,因而在抗肿瘤、抗细胞内寄生感染、移植排斥等功能中起重要作用。
三、造血细胞刺激剂
从多能造血干细胞到成熟免疫细胞的分化发育漫长道路中,几乎每一阶段都需要有细胞因子的参与。最初研究造血干细胞是从软琼脂的半固体培养基开始的,在这种培养基中,造血干细胞分化增殖产生的大量子代细胞由于不能扩散而形成细胞簇,称之为集落,而一些刺激造血干细胞的细胞因子可明显刺激这些集落的数量和大小因而命名为集落刺激因子(CSF)。根据它们刺激的造血细胞种类不同有不同的命名,如GM-CSF、G-CSF、M-CSF、multi-CSF(IL-3)等。目前的研究表明,CSF和IL-3是作用于粒细胞系造血细胞,M-CSF作用于单核系造血细胞,此外Epo作用于红系造血细胞,IL-7作用于淋巴系造血细胞,IL-6、IL-11作用于巨核造血细胞等等。由此构成了细胞因子对造血系统的庞大控制网络。某种细胞因子缺陷就可能导致相应细胞的缺陷,如肾性贫血病人的发病就是肾产生Epo的缺陷所致,正因如此,应用Epo治疗这一疾病收到非常好的效果。目前多种刺激造血的细胞因子已成功地用于临床血液病,有非常好的发展前景。
四、炎症反应的促进剂
炎症是机体对外来刺激产生的一种病理反应过程,症状表现为局部的红肿热痛,病理检查可发现有大量炎症细胞如粒细胞、巨噬细胞的局部浸润和组织坏死,在这一过程中,一些细胞因子起到重要的促进作用,如IL-1、IL-6、IL-8、TNFα等可促进炎症细胞的聚集、活化和炎症介质的释放,可直接刺激发热中枢引起全身发烧,IL-8同时还可趋化中性粒细胞到炎症部位,加重炎症症状.在许多炎症性疾病中都可检测到上述细胞因子的水平升高.用某些细胞因子给动物注射,可直接诱导某些炎症现象,这些实验充分证明细胞因子在炎症过程中的重要作用.基于上述理论研究结果,目前已开始利用细胞因子抑制剂治疗炎症性疾病,例如利用IL-1的受体拮抗剂(IL-1receptor antagonist,IL-lra)和抗TNFα抗体治疗败血性休克、类风湿关节炎等,已收到初步疗效。
五、其它
许多细胞因子除参与免疫系统的调节效应功能外,还参与非免疫系统的一些功能。例如IL-8具有促进新生血管形成的作用;M-CSF可降低血胆固醇IL-1刺激破骨细胞、软骨细胞的生长;IL-6促进肝细胞产生急性期蛋白等。这些作用为免疫系统与其它系统之间的相互调节提供了新的证据。
第四节 细胞因子的临床意义
一、细胞因子与疾病
正常情况下,细胞因子表达和分泌受机体严格的调控,在病理状态下、细胞因子会出现异常性表达,表现为细胞因子及其受体的缺陷,细胞因子表达过高,以及可溶性细胞因受体的水平增加等。
(一)细胞因子及其受体的缺陷
包括先天性缺陷和继发性缺陷两种病理情况,例如先天性的性联重症联合免疫缺陷病人(XSCID),表现为体液免疫和细胞免疫的双重缺陷,出生后必须在无菌罩中生活,往往在幼儿期因感染而夭折。现已发现这种患者的IL-2受体γ链缺陷,由此导致IL-2、IL-4和IL-7的功能障碍,使免疫功能严重受损。细胞因子的继发性缺陷往往发生在感染、肿瘤等疾病以后,如人类免疫缺陷病毒(HIV)感染并破坏TH后,可导致TH细胞产生的各种细胞因子缺陷,免疫功能全面下降,从而表现出获得性免疫缺陷综合征(AIDS)的一系列症状。
(二)细胞因子表达过高
在炎症、自身免疫病、变态反应、休克等疾病时,某些细胞因子的表达量可成百上千倍地增加,例如为风湿关节炎的滑膜液中可发现IL-1、IL-6、IL-8水平明显高于正常人,而这些细胞因子均可促进炎症过程,使病情加重。应用细胞因子的抑制剂有可能治疗这为类症性细胞因子水平升高的疾病。
(三)可溶性细胞因子受体水平升高
细胞膜表面的细胞因子受体可脱落下来,成为可溶性细胞因子受体,存在于体液和血清中,在某些疾病条件下,可出现可溶性细胞因子受体的水平升高。这类分子可能结合细胞因子,使其不再与膜表面的细胞因子受体结合,因而封闭了细胞因子的功能。
二、细胞因子与治疗
目前,利用基因工程技术生产的重组细胞因子做为生物应答调节剂(BRM)治疗肿瘤、造血障碍、感染等已收到良好的疗效,成为新一代的药物。重组细胞因子做为药物具有很多优越之处。例如细胞因子为人体自身成分,可调节机体的生理过程和提高免疫功能,很低剂量即可发挥作用,因而疗效显着,副作用小,是一种全新的生物制剂,已成为某些疑难病症不可缺少的治疗手段。目前已批准生产的细胞因子药物包括干扰素α、β、γ,Epo,GM-CSF,G-CSF,IL-2,正在进行临床试验的包括IL-1、3、4、6、11,M-CSF,SCF,TGF-β等(表4-3、4-4。)这些细胞因子的主要适应症包括肿瘤、感染(如肝炎、AIDS)、造血功能障碍、创伤、炎症等。
表4-3 已批准生产的细胞因子多肽药物
| 药物名称 |
适应症 |
| IFNα |
白血病、Kaposi肉瘤、肝炎、恶性肿、AIDS |
| IFNV |
慢性肉芽、生殖器疣、恶性肿瘤、过敏性皮炎、感染性疾病、类风湿关节炎 |
| G-CSF |
自身骨髓移植、化疗导致的粒细胞减少症、AIDS、白血病、再生障碍性贫血 |
| GM-CSF |
自身骨髓移植、化疗导致的血细胞减少症、AIDS、再生障碍性贫血、MDS |
| Epo |
慢性肾功能衰竭导致的贫血、恶性肿瘤或化疗导致的贫血、失血后贫血 |
| IFNβ |
多发性硬化症 |
表4-4 已批准临床试验的细胞因子多肽药物
| IL-Iα |
放疗、化疗所致的骨髓抑制、恶性肿瘤 |
| IL-1β |
放化疗所致的骨髓抑制、癌症、促进伤口愈合 |
| IL-3 |
骨髓衰竭、血小板缺乏、自身骨髓移植、化疗佐剂、外周血干细胞移植 |
| IL-4 |
免疫缺陷、恶习性肿瘤、疫苗佐剂 |
| IL-6 |
放化疗所致血小板减小、恶习性肿瘤、疫苗佐剂 |
| M-CSF |
恶性肿瘤、白血病、骨髓移植、降胆固醇 |
| TNF |
恶性肿瘤 |
| 干细胞因子(SCF) |
骨髓衰竭 |
| TGF-β |
炎症 |
| IL-11 |
血小板减少症 |
| IL-1受体拮抗剂 |
败血性休克,类风湿关节炎 |
| PLXY321 |
骨髓衰竭 |
细胞因子疗法(cytokine therapy)基本上可分为两种,即细胞因子补充和添加疗法及细胞因子阻断和拮抗疗法。
(一)细胞因子补充和添加疗法
通过各种途径使患者体内细胞因子水平增加,充分发挥细胞因子的生物学作用,从而抗御和治疗疾病。目前已有多种细胞因子(多为基因重组产品)试用于临床治疗,经大量临床资料验证,以下几种细胞因子的临床适应症比较明确,临床疗效比较肯定。
1.IFN 不同型别的IFN各有其独特的性质和生物学活性,其临床应用适应症和疗效有所不同。IFN-α主要用于治疗病毒性感染和肿瘤。IFN-α对于病毒性肝炎(主要是慢性活动性肝炎)、疱疹性角膜炎、带状疱疹、慢性宫颈炎等有较好疗效。IFN-α对于血液系统恶性疾病如毛细胞白血病(有效率达80%以上)等疗效较显著,但对实体肿瘤的疗效较差。虽然IFN-γ的免疫调节作用强于IFN-α,但其治疗肿瘤的效果弱于IFN-α,目前有人应用IFN-γ治疗类风湿关节炎、慢性肉芽肿取得了一定疗效。
2.IL-2 目前多将IL-2与LAD/TIL合用治疗实体肿瘤,对肾细胞癌、黑色素瘤、非何杰金淋巴瘤、结肠直肠癌有较显著的疗效,应用IL-2(或与IFN合用)治疗感染疾病亦取得了一定疗效。
3.TNf 由于其全身应用副作用严重且疗效差,目前多倾向将其局部应用如瘤灶内注射治疗某些肿瘤和直肠癌,其确切疗效尚待进一步评价。
4.CSF 目前主要应用GM-CSF和G-CSF治疗各种粒细胞低下患者。例如与化疗药物合用治疗肿瘤可以降低化疗后粒细胞减少程度,使粒细胞的数量和功能能尽快回升并能提高机体对化疗药物的耐受剂量,从而提高治疗肿瘤的效果。对再生障碍性贫血和AIDS亦有肯定疗效。用于骨髓移植后可使中性粒细胞尽快恢复,降低感染率。此外,应用EPO治疗肾性贫血取得了非常显著的疗效。
(二)细胞因子阻断和拮抗疗法
其基本原理是抑制细胞因子的产生和阻断细胞因子与其相应受体的结合及受体后信号传导过程,使细胞因子的病理性作用难以发挥。该疗法适用于自身免疫性病、移植排序反应、感染性休克等的治疗。例如抗TNF单克隆抗体可以减轻甚至阻断感染性休克的发生,IL-1受体拮抗剂对于炎症、自身免疫性疾病等具有较好的治疗效果。
三、细胞因子的检测
细胞因子检测是判断机体免疫功能的一个重要指标,因而具有重要的实验室研究价值,同时还可能在临床上有诸多实用价值、包括许多疾病的诊断、病程观察、疗效判断及细胞因子治疗监测等。但是,由于细胞因子在体内的含量甚微,给细胞因子的检测带来困维。目前细胞因子的主要检测方法包括:
(一)依赖性细胞株
一些肿瘤细胞株必须依赖于细胞因子方能在体外增殖,如DTLL细胞株依赖IL-2;FDC-PL细胞株依赖于小鼠IL-3;TF-1细胞株依赖于人IL-3和人GM-CSF,因而可利用这些依赖细胞株检测相应的细胞因子。这种方法敏感性高,特异性也不错,但可异的是并非所有细胞因都能找到相应的细胞株,因而限制了它的应用。
(二)功能检测
利用一些细胞因子的功能特性,可建立相应的活性测定方法,如干扰素的抑制病毒感染效应,肿瘤坏死因子对L929细胞的杀伤作用等。这样的方法敏感性高,但特异性不够,容易受一些扰因素的影响。
(三)免疫测定
利用抗原抗体反应的原理,制备出抗细胞因子的单克隆抗体或多克隆抗体,可进行细胞因子的免疫检测。这种方法的优点是特异性强、操作简便,缺点是灵敏度不够,且不能代表活性测定的结果。从目前的国际发展趋势来看,已研制出了高灵敏度、特异性高、高度配套的细胞检测试剂盒,其应用范围正在扩大,有良好的发展前景。
(四)功能测定与抗体抑制
为解决功能定特异性不够,免疫测定灵敏度不够的问题,可将两种方法结合起来,利用各自的长处,有可能得到较为可靠的结果。在这一方法中,所用的抗细胞因子抗体必须是具有中和活性的抗体。
(五)分子杂交技术
利用分子生物学技术,制备出细胞因子的基因探针,可通过分子杂交技术检测细胞内细胞因子mRNA的表达,这是一种高度敏感和高度特异的检测技术,目前在实验室研究中使用较广,其缺点是操作较为繁琐,测定结果只能代表细胞因子基因的表达,而不能代表活性细胞因子的水平。
(六)多聚酶链反应技术(PCR)
PCR(polymerasechain reation)技术是一种高效的基因体外扩增技术,目前PCR技术已用于细胞因子的检测中,首先将细胞因子产生细胞的RNA提取出来,再经逆转录合成cDNA,以cDNA为模板,在细胞因子引物的引导下,即可进行PCR扩增。这种技术是迄今最敏感的细胞因子检测技术,操作也较简便,缺点与分子杂交技术类似,并且不容易对细胞因子表达水平进行定量。

 中医世家
中医世家 浦 标 网
浦 标 网 河南大学精品课程
河南大学精品课程 图书资料室
图书资料室