第一节 抗原的概念和特性
一、抗原的概念
在免疫学发展的早期,人们应用细菌或其外毒素给动物注射,经一定时期后,用体外实验证明,在其血清中存在一种能使细菌发生特异性凝集反应的物质。称之为凝集素,或能特异性中和外毒素性的物质,称之为抗毒素。其后将血清中这种具有特异性反应的物质统称为抗体(antibody,Ab)。而将能刺激机体的物质统称为抗原(antigen,Ag)。
随着现代免疫学的发展,已经证明,当抗原分子进入机体后,能触发免疫细胞一系列的复杂的生物学过程,称之为免疫应答。既能诱导正免疫应答,也能诱导负免疫应答。因此,上述抗原的概念已不能完全概括其函义。目前认为凡能诱导免疫系统发生免疫应答,并能与其产生的抗体或效应细胞在体内或体外发生特异性反应的物质,可称之为抗原。
二、抗原的特性
根据现代抗原的概念,抗原必须具备二种特性,即免疫原性和抗原性。
(一)免疫原性
抗原的免疫原性(immunogenicity)是指抗原分子能诱导免疫应答的特性。它涉及抗原分子与免疫细胞间的相互作用,即它必须经过抗原呈递细胞的加工、处理和呈递作用,以及能被T和B细胞的抗原识别受体所识别。因此抗原的免疫原性与抗原分子的化学性质相关,更与机体的免疫应答特性相关。
(二)抗原性
抗原的抗原性(antigenicity)是指抗原分子能与免疫应答产物,即抗体或效应T细胞发生特异反应的特性,故亦称之为抗原的反应原性(reactivity)。它只涉及抗原分子与抗体分子或T细胞的抗原受体分子(TCR)间的相互作用,即分子与分子间的相互作用。只是抗原分子表面的有限部位能与抗体分子结合,称此部位为抗原决定簇(antigen determinant)或表位(epitope)。因此抗原的抗原性主要决定于抗原分子的化学性质。如抗原为蛋白质分子,其抗原性可决定于其氨基酸序列或其空间构型。
第二节 抗原的免疫原性
抗原的免疫原性,首先决定于其自身的化学特性,但同一种抗原,对不同种动物或同种动物不同个体间其免疫原性强弱,可表现很大差异,因此一种抗原的免疫原性是由其化学性质和宿主因素决定的。具有免疫原性的物质称为免疫原(immunogen)。
一、免疫原性的化学基础
许多天然物质可诱导免疫应答,其中大分子蛋白质和多糖具有强免疫原性,小分子多肽及核酸也具有免疫原性。
(一)化学组成
大分子蛋白质,分子量大于10000者,可含有大量不同的抗原决定簇,是最强的免疫原。如异种血清蛋白、酶蛋白及细菌毒等,是强免疫原蛋白质的例子。
多糖是重要的天然抗原,纯化多糖或糖蛋白、脂蛋白以及糖脂蛋白等复合物中的糖分子部分都具有免疫原性。在自然界,许多微生物有富含多糖的荚膜或胞壁,细菌内毒素是脂多糖,以及一些血型抗原(A、B、C、H)也是多糖。
核酸分子多无免疫原性,但如与蛋白质结合形成为核蛋白则有免疫原性。在自身免疫病中,可见对天然核蛋白诱导的免疫应答产生的抗DNA或RNA抗体。
此外,多肽类激素如胰岛素虽为小分子量(6000)亦具有免疫原性。来自一种动物的胰岛素,如长期用于另一种动物,亦能诱导免疫应答产生抗体。
(二)分子量
凡具有免疫原性的物质,其分子量都较大,一般在1万以上,小于1万者呈弱免疫原性,低于4000者一般不具有免疫原性。许多小的免疫性原性分子可激发细胞免疫,而不产生抗体。亦有大分子量物质,如明胶分子量可达10万,但因其为直链氨基酸结构,易在体内降解为低分子物质,所以呈弱免疫原性。可见免疫原性除与分子量有关外,还与其化学结构相关。
(三)化学结构
在蛋白质分子中,凡含有大量芳香族氨基酸,尤其是含有酷氨酸的蛋白质,其免疫原性强;而以非芳香族氨基酸为主的蛋白质,其免疫原性较弱。蛋白质和多糖抗原,凡结构复杂者免疫原性强,反之则较弱。其复杂性是由氨基酸和单糖的类型及数量等决定的。如聚合体蛋白质分子较单体可溶性蛋白质分子的免疫原性强。
二、宿主因素与免疫原性
免疫原性物质进入机体后能否诱导产生免疫应答,除上述化学基础外,尚受宿主因素的影响,而且是更要的因素。
(一)异种性
在免疫功能正常条件下,只有异种或同种异体的免疫原性物质才能诱导宿主的正免疫应答,即只有“非已”抗原才能引起正免疫应答。这是由于免疫系统在个体发育过程中,对“自己”抗原产生耐受,不能识别,而对“非已”抗原则能够识别所致。因此抗原来源与宿主种系关系越远,其免疫原性也越强,如微生物抗原、异种血清蛋白等物质对人是强免疫原。反之种系关系较近,则免疫原性也弱,如鸭血清蛋白对鸡呈弱免疫原性,而对兔则表现为强免疫原性。得但目前证明在正常个体也可诱发生理性自身免疫应答,只有超出一定范围才能直病理性自身免疫应答。
(二)宿主遗传性
在同种动物不同个体间结感染的抵抗力存在明显差异的事实,已早为人知,但不能解释其产生的原因,只以个体差异说明之。其后,用已知人工合成抗原给不同近交系动物免疫,每一近交系动物其遗传背景相同,结果发现有的品系能产生抗体,称为高应答品系(hkgh responder)。有的品系不能产生抗体,称为无或低应答品系(nonrespondre)。例如应用人工合成抗原二硝基苯-多聚-左旋-赖氨酸(DNP-poly-L-L)在荷兰猪品系2(GP strain2)可以引起应答,而对品系13(GP strain13)则不能引起应答。这充分证明个体遗传性对免疫应答的控制作用。在70年代McDevitt等应用人工合成抗原在近交系小鼠体内发现了控制免疫应答的基因座(immuneresponslocus)定位于H-2复合体的Ⅰ区,称此基因为免疫应答基因-1(immune uesponse,Ⅰr-1)。
(三)免疫佐剂
免疫佐剂的物质有:①微生物及其产物,常用的微生物有分枝杆菌、短小棒状杆菌、百日咳杆菌以及处左兰氏阴性杆菌的提取物脂多糖,自分枝杆菌的提取物质胞壁酰二肽等。②多聚核苷酸,如多聚肌苷酸:胞苷酸(poly1:C),多聚腺苷酸(poly1:A:μ)等。③弗氏佐剂(Freundadjuvant),这是目前最常用于动物实验的佐剂,它是将抗原水溶液与油剂(石蜡油或植物油)等量混合,再加乳化剂(羊毛脂或叶吐温80)制成油包水抗原乳剂,称之为不完全弗氏佐剂。如在不完全佐剂中加入分枝杆菌(如死卡苗)则称为完全弗氏佐剂。④无机物,如明矾及氢化铝等。
佐剂的作用机制尚不完全清楚:①它可能增加抗原的表面面积,易为巨噬细胞所吞噬;②延长抗原在体内的存留期,增加与免疫细胞接触的机遇;③诱发抗原注射部位及其局部淋巴结的炎症反应,有利于刺激免疫细胞的增殖作用。
第三节 抗原的抗原性
自发现抗体后,用血清学方法在体外实验,证明了天然抗原与其相应抗体发生特异性结合,这是一个重要的免疫学现象,称这种特性为抗原的抗原性。在早期由于尚未建立对蛋白质抗原进行分析的方法,为研究抗原性的化学本质造成了困难。奥地利免疫化学家Landsteiner在本世纪20年代创建了人工结合抗原,并应用血清学方法对抗原性的化学本质进行了系统的研究,为抗原、抗体的相互作用提供了大量知识,为天然抗原化学性质的研究奠定了基础。
一、人工结合抗原
Landsteiner首创用已知化学结构的低分子化合物或化学基团与免疫原性蛋白质进行偶联,制备了偶氨蛋白。由于低分子化合物本身无免疫原性,单独免疫动物不能对其产生抗体,如与蛋白质偶联再免疫动物则能对其产生抗体,并能单独与其抗体结合。故称此种无免疫原性低分子化合物为半抗原(hapten,H),称免疫原性蛋白质为载体(cauuier,C)自此建立了半抗原与载体的概念(H-C)。因此可以认为半抗原是外加在免疫原分子上的已知化学结构的化学基团,在一定意义上它与抗原分子上未知化学结构的天然抗原决定簇是同义语(图10-1)。
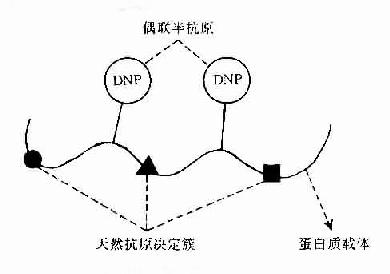
图10-1 半抗原载体结合物
常用为载体的免疫原性蛋白质有牛血清蛋白(BSA)、牛血清丙种球蛋白(BGG)、卵清蛋白(OA)、鸡丙种球蛋白(CGG)、钥孔嘁血蓝蛋白(KLH)及人工合成多聚赖氨酸(PLL)等。常用为已知结构的半抗原有二硝基苯酚(DNP)和三硝基苯酚等低分子化分物。
Landsteiner有带有不同酸基的有机物为半抗原,分别与同一种载体偶联制备了几种酸基不同的半抗原载体结合物,然后免疫动物。用所得抗体与已知半抗原进行体外补体结合实验,结果证明,带不同酸基的半抗原只能与其相应抗体结合。同理,他用氨基苯甲酸的三种异构物(邻位、间位、对位)分别与同一种载体蛋白偶联的半抗原载体结合物,只能与其相应抗体结合(表10-1,2)。上述实验证明,抗原与抗体的特异结合是与抗原分子表面的特殊结构的化学基团相关。将抗原分子表面能与其抗体结合的部位称为抗原决定簇,即抗原的抗原性是由其抗原决定簇的性质、数目和空间构型所决定,自此建立了抗原决定簇的概念。为以后研究天然蛋白质的抗原性提供了理论依据。
表10-1 不同酸基对半抗原与抗体反应特异性的影响
|
半抗原 |
| 抗下列半抗原-载体 免疫血清 |
苯胺NH2 |
对氨基苯甲酸NH2 |
P对氨基苯磺酸 |
对氨基苯砷酸 |
| 载体-苯胺 载体-对氨基苯甲酸 载体-对氨基苯磺酸 载体-对氨基苯砷酸 |
+++ - - - |
- ++++ - - |
- - ++++ - |
- - - +++ |
表10-2 化学基团(COOH)位置对半抗原体反应特异性的影响
|
半抗原 |
| 抗下列半抗原-载体 免疫血清 |
苯胺 |
氨基苯甲酸 |
| 邻位 |
间位 |
对位 |
| 载体-苯胺 载体-邻位氨基苯甲酸 载体-间位氨基苯磺酸 载体-对位氨基苯砷酸 |
+++ - - - |
- ++++ - - |
- - ++++ - |
- - - +++ |
+:阳性反应;-:阴性反应
抗原除与其相应抗体发生特异反应外,也可与其它相关抗体发生反应,称这种现象为交叉反应。这是由于在二种不同的抗原分子中具有相同的抗原决定簇引起的,称为共同簇或共同抗原.也可能由于二种不同决定簇具有相似的结构,但并非相同,也能引起交叉反应。在发生学上较近的种属间,其抗原产生的抗体,彼此间易出现交叉反应。如牛血清清蛋白(BSA)与其它种属清蛋白之间的交叉反应(表10-3)。
表10-3 牛血清蛋白与其它种属清蛋白的交叉反应
| 清蛋白来源 |
交叉反应%(BSA) |
清蛋白来源 |
交叉反应 %(BSA) |
| 人 猪 羊 马 地鼠 |
15 32 75 13 13 |
豚鼠 狗 小鼠 大鼠 猫 |
5 13 10 17 25 |
二、载体决定簇与半抗原决定簇
单独应用半抗原不能诱导机体产生抗体,只有将半抗原与载体蛋白结合后,才能诱导机体既能产生抗载体蛋白抗体,也能产生半抗原抗体,已为实验所证明。这就提出了为什么单独应用半抗原不能产生抗体,载体在抗体产生中发挥什么作用等问题。
为了回答上述问题,有的学者应用牛血清蛋白-二销基苯(BSA-DNP)和卵白蛋白-二硝基苯(OA-DNP)为载体结合半抗原,观察了载体在抗体产生中的作用,他们首先应用BSA-DNP对小鼠进行初次免疫,然后再分别应用BSA-DNP和OA-DNP进行再次免疫,观察载体对抗DNP抗体产生的影响.他们发现,只有当初次与再次免疫时,半抗原需要在相同载体上才能产生半抗原抗体,称此为载体效应(表10-4)。证明载体不是单纯起运载半抗原的作用,而是具有载体特异性。因此提出一个完全抗原分子,必须具有载体决定簇和半抗原决定簇。
表10-4 载体-半抗原效应
| 实验组别 |
初次免疫 |
再次免疫 |
抗DNP抗体 |
| 1 2 3 |
BSA-DNP BSA-DNP BSA-DNP+OA |
BSA-DNP OA-DNP OA-DNP |
+++ + +++ |
其后Mitchison等在70年代应用载体效应过继转移实验,证明了在抗体形成过程中,有对载体特异的细胞和对半抗原特异的细胞,分别称为载体反应细胞和半抗原反应细胞。并进一步证明T细胞是载体反应细胞,对抗体产生起辅助作用。B细胞是半抗原反应细胞,是产生抗体的细胞,自此阐明了载体效应的细胞学基础。
载体-半抗原概念极其重要,它可解释为什么低分子量化合物与体内载体蛋白质分子结合诱发超敏性反应产生的药物过敏症。如苯胺类染料、镇静剂司眠脲、退热剂阿司匹林、氨基比林以及多种抗生素分解产物等是诱发药物过敏症的原因。
三、天然蛋白质的抗原性
绝大多数免疫原性物质是大分子蛋白质或细胞组分,但只是其有限的部位能与其相应抗体结合,此部位称为抗原决定簇或表位(前述载体-半抗原效应已证明一个抗原分子须有T细胞决定簇和B细胞决定簇)。由于天然大分子蛋白质结构复杂,并具有空间构型,因此对其单一抗原决定簇的化学组成、定位及其结构研究十分困难。但近年来由于免疫化学技术的发展,已能研究其空间结构,由于单克降抗体技术的发展,已能分离鉴定单一抗原决定簇,因此,对蛋白质抗原性的研究已取得很大进展。
(一)B细胞决定簇(表位)
抗原分子的B细胞决定簇大小不同,其最大表面积约为50~70mm约由4~6氨基酸残基或糖基组成。100个氨基酸残基多肽可有14到20个非重叠决定簇,由线状排列彼此相邻的氨基酸组成,故称为线性或连续性决定簇。而球蛋白是有三维空间的折叠肽链,故其大多数决定簇被掩盖在内部,可称为隐蔽性决定簇。只存在于其表面的决定簇可被免疫细胞识别,或与抗体结合者称为功能性决定簇(图10-2)。组成这种决定簇的氨基酸,是由折叠的肽链将有同位置的氨基酸使之相邻成为有一定空间构型的决定簇,故称为构像决定簇或不连续决定簇。现已证明抗原分子的抗原性是由其组成的氨基酸予列、空间构型及其决定簇片段的移动决定的。
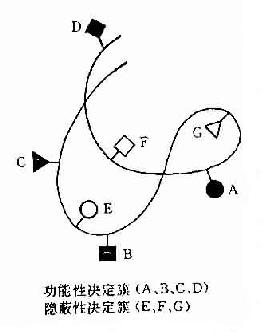
图10-2 球蛋白分子抗原决定簇
在球蛋白分子表面可带有多种抗原决定簇,其中有些决定簇更容易与淋巴细胞接触,比其它决定簇能激发较强的免疫应答,称这种决定簇为优势决定簇。每个抗原分子上其功能性决定簇的总数称为抗原价,即能与抗体分子起反应的最大数量的抗原决定簇,一般分子量大于10万者可有2个抗原价,大分子如分子量为40万者可有10~12个抗原价。
(二)T细胞决定簇(表位)
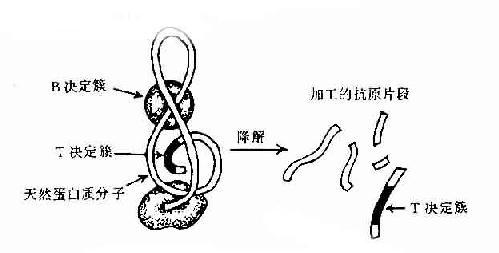
图10-3 抗原分子的T与B决定簇
现已证明T细胞决定簇为免疫原性多肽片段,属连续性决定簇。但它不存在天然蛋白分子表面,必须经抗原呈递细胞加工处理为小肽分子,然后再与自己MHC分子结合才能被T细胞所识别。而B细胞决定簇可存在于天然抗原分子表面,不须经加工处理,即可直接被B细胞所识别(图10-3)。
在天然大分子蛋白质抗原直接证明其存在有T细胞决定簇和B细胞决定簇是十分困难的。但用小分子已知结构的免疫原进行研究已证明其存有2种决定簇。胰腺高血糖素激素(glucagon)由29个氨基酸组成,有免疫原性。由其产生的抗激素抗体可识别其氨基末端。由其产生的杀伤T细胞可识别其羧基末端,相当于激素分子的载体部分。由此证明胰高血糖素分子的氨基端为B细胞决定簇,而羟基端为T细胞决定簇(图10-4)。
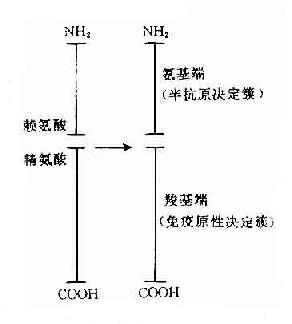
图10-4 高血糖素决定簇结构示意图
第四节 抗原的分类
一、天然抗原
根据抗原性物质与机体的亲缘关系可分为“自己”(self)与“非已”(non-self)抗原。即与机体种系发生关系愈远,其遗传性差异越大,其免疫原性也愈强。
(一)“自己”抗原
正常自身组织成分及体液组分处于免疫耐受状态,不能激发免疫应答,但如打破自身耐受,则可引起自身免疫应答;另一些自身组织成分虽具有免疫原性,但在正常情况下,由于组织屏障,不能进入血流,因此不能与免疫细胞接触,也不能激发免疫应答,称此种抗原为隐蔽性身抗原,如脑组织、眼晶状体蛋白及精子等。一旦因外伤或手术等原因,可使此种抗原进入血流时,则可引起自身免疫应答。受病原微生物的感染或应用某些化学药物,可与自身组织蛋白结合,改变其分子结构,形成修饰的自身抗原,也能引起免疫应答。
(二)非已抗原
来自异种动物的抗原物质称为异种抗原。如来自外部侵入人体的各种病原微生物及其产物的外毒素,注射的异种动物免疫血清,以及吸入和食进的异种蛋白,例如花粉和食物均属异种抗原。由于与人种属关系远为强免疫原。此外癌细胞可在人体内产生特异性癌抗原,但对其免疫原性迄今尚未能证实。
在同种动物不同个体间也存在各种组织成分抗原性的差异,称此种抗原为同种异型抗原。这种抗原受遗传支配,它可在遗传性不同的另一些个体内引起免疫应答,称之为异型免疫应答。如人血型抗原不同输血时可引起输血反应,组织相容性抗原或移植抗原型不同可引起移植排斥反应。此外,免疫球蛋白分子上存在的Gm、Am、Km标记均属异型抗原,可用以鉴别IgG、IgA及K轻链的异型。
在不同种属动物组织间也可发现有共同抗原,称这种抗原为异嗜性抗原。Forssman首先发现这种抗原,故亦称之为Forssman抗原。即这种抗原无种属特异性,它可共同存于人、不同种动物与微生物之间,因此它与疾病的发病学和诊断有一定意义。
目前已发现多种异嗜性抗原,如大肠杆轿O86含有人的B血型物质,肺炎球菌14型含有人A血型物质,它们与人血型抗体的产生有关。有些病原微生物与人体某些组织具有共同抗原成分,是引起免疫性疾病的原因之一。如溶血性链球菌一些抗原可与肾小球基底及心肌组织有有共同抗原成分,它们可能与急性肾小球肾炎和风湿病的发病有关。又如大肠杆菌O14型脂多糖与人结肠粘膜有共同抗原,可能与溃疡性结肠炎的发病有关。
某些疾病的诊断也可借助于对异嗜性抗原的检测。例如引起原发性非典型肺炎的病原支原体与MG株链球菌有共同抗原,可藉其血清中抗体对此种链球菌的凝集反应进行诊断,引起斑疹伤寒的立克次体与变形杆菌一些菌株间有共同抗原,可藉其血清中抗休对变型杆菌的的凝集反应进行诊断,称之为Weil-Felix反应。此外传染性单核细胞增多症患者血清中,可出现凝集羊红细胞的异嗜性抗体,可用羊红细胞凝集反应进行诊断。
二、人工抗原
用化学合成法或基因重组法制备含有已知化学结构决定簇的抗原,称之为人工抗原。它可包括人工结合抗原、人工合成抗原和基因重组抗原。无论对免疫学理论研究和分子疫苗的制备都具有重要意义。
(一)人工结合抗原
将无免疫原性的简单化学基团与蛋白质载体偶联,或将无免疫原性的有机分子如二硝基苯(DNP)或三硝基苯(TNP)与蛋白质载体结合,形成载体-半抗原结合物,均属人工结合抗原。应用此种抗原证明了抗原与抗体特异结合的化学基础,以及在抗体生成过程中T与B细胞的协同作用。
(二)人工合成抗原
用化学方法将活化氨基酸聚合,使之成为合成多肽,只由一种氨基酸形成的聚合体称为同聚多肽,如由左旋赖氨酸形成的共同聚多肽(PLL)。由二种或二种以上氨基酸形成的聚合多肽称为共聚多肽,如由酪氨酸、谷氨酸与多聚丙氨酸和赖氨酸组成的聚合成多肽(T、G)-AL。应用这种人工合成多肽可研究氨基酸种类、序列与蛋白质抗原性及免疫原性的关系,也可研究机体遗传性与免疫性的关系。
对天然蛋白质抗原性的研究证明,任何一个氨基酸片段,只要具有合适的构型,都有抗原性,甚至一小段合成的小肽与合适的载体相联接,也能诱导产生抗体,并能与其天然分子构型相结合,这就提示,可根据天然蛋白质抗原的免疫原性片段进行氨基酸序列分析,或由其编码DNA推导的氨基酸序列,进行构建人工合成多肽疫苗。
(三)基因工程抗原
近年来由于分子生物学技术的进步,已有可能将编码免疫原性氨基酸序列的基因克隆化并与适当载体(如细菌粒或病毒)DNA分子相结合,然后引入受体细胞中(如原核细胞的大肠杆菌或真核细胞酵母菌及哺乳类动物细胞)使之表达,即能获得免疫原性之融合蛋白,经纯化后可做为疫苗,此即基因工程疫苗。
应用分子生物学技术制备基因重组疫苗的另一进展,是将目的抗原决定簇的DNA序列插入另一种比较安全的活病素基因组中(如牛痘苗),制备所谓重组感染载体多价疫苗。
随着70年代分子病毒学的发展,特别是对病毒基因的结构、功能与复制方面知识的积累,为迅速研制病毒亚单位疫苗、合成多肽苗以及基因工程疫苗奠定了基础。
一些重要病毒如乙型肝炎病毒、脊髓灰质炎病毒、疱疹病毒以及流感病毒等的蛋白质多肽,都已利用基因工程进行了成功的表达,有的已进入临床试验阶段。我国也报导了正在进行研制基因工程乙型肝炎病毒疫苗和在牛痘苗表达系统中研制乙肝病毒的重组感染载体的多价疫苗。
三、胸腺依赖抗原与胸腺非依赖抗原
实验证明,由抗原激发的免疫应答是多细胞相互作用的结果,即由抗原呈递细胞、T细胞和B细胞共同参与予完成的。大多数抗原激发的体液免疫应答,必须有TH细胞参予才能完成,称这种抗原为胸腺依赖抗原(thymus-dependent antigen,TD Ag)。但也有少数抗原物质,不须TH细胞参予,可单独刺激B细胞产生抗体,称这种抗原为胸腺非依赖抗原(thymus independentantigen,TIAg)。这二种抗原的区别主要在于其抗原决定簇的结构不同所致。TD抗原在其分子结构上,既具有载体功能的决定簇,也具有抗原性决定簇。且在其分子表面出现多种不同抗原决定簇,但缺乏重复出现的同一决定簇,TD抗原主要是大分子蛋白质。而TI抗原多数为大分子多聚体,带有重复出现的同一抗原决定簇,且降解缓慢,故不须TH参加即能直接刺激B细胞,TI抗原主要是多糖类物质(图10-5、表10-5、6)。
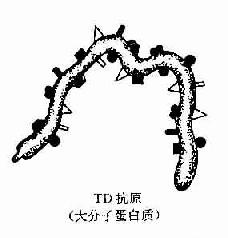
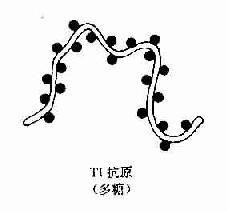
图10-5 TD与T1抗原种类
表10-5 TD与TI抗原种类
| TD |
TI |
| 人丙种球蛋白 牛血清清蛋白 卵白蛋白 类毒素 |
羊红细胞 组织相容性抗原等 |
肺炎球菌荚膜多糖 脂多糖 聚合鞭毛 DNP-聚蔗糖 聚乙烯基吡络烷酮(PVP) |
表10-6 TD抗原与TI抗原的特性
| 特性 |
TD抗原 |
TI抗原 TⅠ-1 TⅠ-2 |
| 化学特性 |
蛋白质 |
脂多糖 |
多糖 |
| 抗体应答 |
|
+ |
+ |
| 无胸腺鼠 |
- |
+ |
-/少 |
| 无T细胞培养物 |
- |
|
|
| 抗体应答的特点 |
- |
|
|
| 类别转换 |
+ |
- |
- |
| 亲和性成熟 |
+ |
- |
- |
| 记忆B细胞 |
+ |
- |
- |
| 多克隆B细胞活化剂作用 |
- |
+ |
- |
| 诱导DTH能力 |
+ |
- |
- |
四、超抗原
(一)超抗原的概念
超抗原(supper antigen,Sag)是一类由细菌外毒素和逆转录病毒蛋白构成的抗原性物质。它们能与多数T细胞结合并为T细胞活化提供信号。而上述的普通抗原只能与少数对应T细胞结合并使之活化。因此称这种能与多数T细胞结合的抗原为超抗原。
(二)超抗原与T细胞结合的特征
超抗原主要与CD4+T细胞结合,而和普通抗原肽与T细胞的结合有很大差异。超抗原既能与APC细胞上MHCⅡ类分子结合,也能与TCR Vβ链结合是其作用特点。
超抗原无需经APC加工可直接与MHCⅡ类分子非多态区外侧结合,而不是与肽结合沟结合,故无MHc 限制性。
在T细胞方面超抗原只与TCRVβ片段结合,而与D和J区无关,也与TCRα链无关。任一已知超抗原能与其特殊殊的Vβ片段结合,所以一种超抗原可活化多数T细胞,约占T细胞库的1/20~1/5,这远远超过普通抗原活化T细胞的数量(表10-7)。
表10-7 超抗原的作用特性
|
普通抗原 |
超抗原 |
| T细胞一次应答 |
- |
+ |
| T细胞反应频率 |
1/106~1/104 |
1/20~1/5 |
| MHCⅡ类分子 |
肽结合沟 |
非多态区(α-螺旋) |
| 结合部位 |
|
外侧 |
| MHC限制性 |
+ |
- |
| APC存在 |
+ |
|
(三)超抗原的种类
1.内源性超抗原(病毒性) 70年代初Festenstein发现在MHC相同,而MHC以外基因区不同的纯系鼠间进行淋巴细胞混合培养,可引起很强的T细胞增殖反应,将刺激这种增殖反应的抗原称为次要淋巴细胞刺激抗原(minor lymph'ocyte stimulating antigen,Mlsag)。
近年来证明这种内源性MLs抗原是小鼠乳腺肿瘤病毒(mouse mamary umor virus,MMTV)产生的蛋白。MMTV是一种逆转录病毒,以前病毒(provirus)形式整合于小鼠细胞DNA中。这种小鼠可终生制造这种病毒蛋白,因之可视为一种自身超抗原。这种小鼠内源性MLs抗原的化学性质现已证明是一种糖蛋白。
由于MLs抗原的来源已经清楚,故目前称这种小鼠的内源性超抗原为病毒性超抗原。人类是否也有这种病毒性超抗原,目前尚不能肯定,但有人提出人类免疫缺损病毒(HIV)也是逆转录病毒,有可能是人类的病毒性超抗原。
2.外源性超抗原(细菌性)外源性超抗原是一类细菌性外毒素组成,主要由革兰氏阳性细菌产生。如金黄色葡萄球菌产生的肠毒素(staphylococcus enterotoxin,SE)以及链球菌产生的致热外毒素等。
(四)超抗原的生物学意义
1.超抗原与T细胞的耐受诱导实验证明在胸腺内分化发育中的T细胞如与超抗原结合,可诱发程序性细胞死亡,导致克隆排除。用抗Vβ单克隆抗体在周围血中检测不出带有特殊Vβ受体的T细胞,为T细胞耐受诱导机制的研究提供了有力的实验模式。
2.超抗原与疾病葡萄球菌感染所产生的外毒素主要是可溶性蛋白分子,近年来的研究证明葡萄球菌外毒素对靶细胞并无直接毒性作用,而是通过活化多数T细胞所释放的大量细胞因子产生的生物学效应引起的毒性休克综合征等临床征状。
一些疾病,例如原因不明的川畸病,风湿性关节等疾病,发现与某些Vβ阳性T细胞的增殖相关。周围组织中存在的自身反应性T细胞克隆可为外源性超抗原激活而引发自身免疫病。也有学者认为HIV引发的人艾滋病,其发病学与其超抗原相关。

图10-6 抗原分类

 中医世家
中医世家 浦 标 网
浦 标 网 河南大学精品课程
河南大学精品课程 图书资料室
图书资料室