第一节 近交和近交系
一、近交
实验动物的近交(Inbreeding)即近亲繁殖,是指血缘关系极为相近的个体之间或遗传组成极相似的个体之间进行的交配繁殖。是培育近交系动物的必须手段,通过近交使一个种群达到接近完全纯合程度,即所有同源染色体的相对位置都具有相同基因的状态。因此,通过同胞兄妹或亲子交配可以较快获得近交品系动物。
从同一祖先获得遗传物质,一条得自父,一条得自母,来源于共同祖先的小动物自交,它们得自父母同基因比非近交者多。近交和非近交繁殖时亲缘关系可见图3-1。
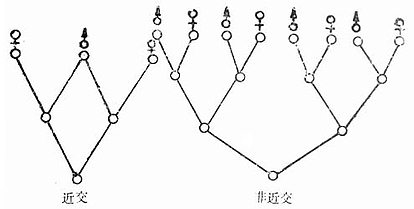
图3-1 近交和非近交时亲缘关系示意图
近交可以降低杂合性,可以将群体分离为不同的品系,然而近交的结果也必将出现近交衰退。
二、近交后引起的变化
(一)近交可以增多纯合性,也就是降低杂合性(Reduction of the terozygosity)
自然界的动植物在遗传上一般都是杂合子体,故最初培育近交系动物都是取源于杂种,换句话说也就是取源于异型接合体动物。动物的遗传纯度取决于交配动物之间遗传关系的远近。因此近交系动物通过长期近亲交配而建立的。通过纯育后,动物的杂合性逐渐降低,因而纯度逐渐增高,使基因位点变为纯合子,使它们的表现型趋向一致性,如A与a基因频率相同,但基因型分配变化,经过一定代数的兄妹交配,则不再有杂合子,而获得遗传稳定的AA与aa近交品系,见表3-1。同胞兄妹或亲子交配连续20代以上就培育成近交系,为了保持该系的遗传特点,必须继续近交,并不得因突变或遗传漂移而丢失。
表3-1 近交引起的基因纯合情况
P(A)=0.5 p(a)=0.5
| 代数 |
各 种 基 因 的 比 例 |
| 纯合子(野鼠色)AA |
杂合子(野鼠色)Aa |
纯合子(黑色)aa |
| 原 始 种 群 |
1/4 |
2/4 |
1/4 |
| 兄妹交配下一代 |
5/16 |
6/16 |
5/16 |
| 兄妹交配下二代 |
11/32 |
10/32 |
11/32 |
| 兄妹交配下三代 |
24/64 |
16/64 |
24/64 |
| 兄妹交配20代以后 |
50/100 |
0 |
50/100 |
实验动物交配繁殖中的一些概念和符号。
1.纯合子:同源染色体的相对位点上具有两个相同的基因称为纯合子(Homozygote),如AA(显性纯合子),aa(隐性纯合子)。
2.杂合子:两个基因不相同时,即由一个显性基因和一隐性基因结合而成的杂合子交(Heterozygote),如Aa。
3.杂合子的a位点:即指a位点上两个位点为A/a。
4.纯合子的等位点:用A/A或a/a表示。
5.显性突变位点用D表示,隐性突变位点用r表示。
6.D和r位点在繁殖制度中被称作重要位点(Loci of interest),因为它们是控制位点分离的。
7.D位点上的等位基因为A/D,r位点上的等位基因为A/r。
8.以在一个位点上的等位基因A/A、a/A、a/a三种基因型进行组合将产生九种交配形式,可归纳为以下四类交配系统。
(1)纯合子交配(Incrosses):为亲代相同的显性纯合子交配,如A/A×A/A和a/a×a/a。
亲代:♂AA×♀AA 子代:基因型AA,表现型A。
(2)杂交(Crosses):为亲代不相同的纯合子交配,如A/A×a/a和a/a×A/A。
亲代:♂AA×♀aa 子代:基因型Aa ,表现型A。
(3)回交(Backcrosses):为亲代显性纯合子与杂合子交配。如A/A×a/A;a/A×A/A;a/A×a/a和a/a×a/A
亲代:♂Aa×♀Aa 子代:基因型:1AA,1Aa 表现型:A,均为显性野鼠色。
(4)自交或称互交(Intercrosses):亲代为两个杂合子交配:如a/A×a/A。
亲代:♂Aa×♀Aa 子代:基因型:1/4AA,2/4Aa,1/4aa。表现型:3/4A,1/4a。
| 精子 卵子 |
A |
a |
| A |
AA |
Aa |
| Aa |
Aa |
Aa |
C57BL黑色a,C3H野鼠色,二者皆为纯合子。二者杂交:

种系中纯合性的程度通常以近交系数(F)表示,所谓近交系(Coefficient of inbreeding)也就是一个种系的个体中,两个等位基因具有相同来源的概率,通常以0~1之间的尺度来表示。F=0为完全杂合,F=1表示完全纯合。F值随近交繁殖连续的代数而递增。增加的比率决定于交配动物亲缘关系的密切程度。根据Feiconer(1960)的研究,认为全同兄弟姐妹交配,近交系前几代数值不恒定,如前四代近交系数上升率分别为28%、17%、20%和19%,以后每代上升率就恒定为19.1%。故Felconer提出一个便于计算的公式:Fn=1-(1-△F)n。n表示近交代数,△F是每进一代的近交系数上升率。例如繁殖了10代,△F为19%,代入公式,则
F10=1-(1-0.19)10=1-(0.81)10=1-0.1216=0.8784=87.84%
也就是说繁殖到第10代时,纯度可达到87.84%,还有12.16%是杂合的。由于交配方式不同,其△F也各不相同,同胞兄妹交配△F为19.1%,同父异母交配11.0%,回交(亲子交配)19.1%,堂兄妹交配为8%。全同胞兄妹或亲子交配前20代的近交系数计算状况可见表3-2。
表3-2 Felconer近亲系数表
| 代 数 |
F |
代 数 |
F |
| 1 |
0.250 |
11 |
0.908 |
| 2 |
0.375 |
12 |
0.926 |
| 3 |
0.500 |
13 |
0.940 |
| 4 |
0.594 |
14 |
0.951 |
| 5 |
0.672 |
15 |
0.961 |
| 6 |
0.734 |
16 |
0.968 |
| 7 |
0.785 |
17 |
0.974 |
| 8 |
0.826 |
18 |
0.979 |
| 9 |
0.859 |
19 |
0.983 |
| 10 |
0.886 |
20 |
0.986 |
注:近交系不改变频率。
(二)近交可将群体分离为不同基因型的品系(Splitting of the Population into Lines of Different Gentypes)
原始种群经过近交分为若干近交系,在不同等位基因的不同位点上变为纯合子。原始种群的小鼠兄妹交配连续20代之后分离出两支等位基因A和a的近交品系:
1.等位基因A的品系:只生产野鼠色后代。
2.等位基因a的品系:只生产黑色后代。
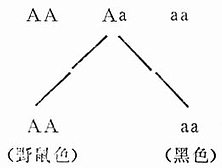
近交品系动物的获得,可使原来杂合子的动物增加纯合性(Homozygosity),从而提高基因型的稳定性,获得想要的品系,实验者可以从动物的特定性状中进行选择。
(三)近交可引起近交衰退(Inbreeding Depression)
亲缘接近的交配所产生的后代常常会出现生长、成活、生育、抗病、适应环境等能力的减退,这种现象称之为近交衰退,如何克服和解决这个问题,是培育近交系动物的关键所在。由于近交衰退的缘故,从一个供繁殖的种群培育和建立一个近交系时,往往需要从杂交群或几个近交系开始才能成功,而且的近交几代后,有些系可以因生育或生活率低而断绝;留下的受近交衰退影响较小的后代有些可以维持下来,而另外的也可能最后被淘汰。也由于同样的原因,过去建立的某些近交系,现在只有某些亚系还仍然存在,有些不能全部存活,甚至难以维持。
近交衰退发生的原因是多方面的,从遗传学的角度解释主要有两点:
1.有害的隐性基因的暴露。一般病态的突变基因绝大多数都是隐性的,所以处于杂合状态时是不表现出病态或不利的性状。这些有害基因的作用可被显性的杂合子等位基因所掩盖,但经过一段近亲繁殖,纯合的基因(纯合子)比例渐渐增多,于是有害的隐性基因相遇成为纯合子而显出作用,出现了不利的性状,对个体的生长发育、生活和生育等产生明显的不利影响。例如杂种动物所带有的不育的隐性基因往往被其显性的等位基因所掩盖,而不表达其不育的性状,但由于纯育,动物的纯合性逐渐增高,不育的现象也就表现出来了。
2.多基因平衡的破坏。个体的发育受多个基因共同作用的影响,虽然其中每个基因的作用效应微小。对环境适应较好的野生或杂交动物,由于自然选择的作用有利于保存那些生物适应能力较强的基因组合具有平衡的多基因系统,近交繁殖往往会破坏这个平衡,造成个体发育的不稳定。
近交衰退往往在近交培养过程中的最初若干世代中表现出来,以后经过一定的人工选择,带有纯合有害基因的动物被逐渐淘汰,或者由于无意识地保留了一小部分的杂合性,经过5~10代左右的培育繁殖,后代中生育与生活力可以逐渐稳定,不再下降。
由于近交系动物是采用近亲繁殖方式,因此容易引起生活力的降低、生长繁殖力退化、抗病力降低等近交衰退变化,为了防止这些问题产生,应注意如下几点:
1.为防止种群传代终断,可采用回交方式(即父×女或母×子)而不计算近交代数,这样可以维持种群不致在繁殖中终断。
2.饲料营养的保证很重要,在一般传代至5~7代时会出现生命力下降,生长繁殖退化,或出现产仔畸形等情况,因此应在饲料中适当增加营养成分以保证子代生长发育正常。
3.在传代中注意检查子代的情况选优去劣,选择体质健康、生殖力旺盛的后代而不能选择体弱繁殖力差的动物。
4.近交到6~7代时,可出现致死的有害基因,而且这种致死基因随近交代数增加而不断升高,可引起个体死亡,为此,培育近交品系,开始不必用同窝兄妹交配,用同父异母,或同母异父的兄妹交配,如待近交品系建立后再改变致死基因则很困难。
第二节 近交系的特征和应用意义
一、近交系的特征
(一)基因纯合性。通过持续20代以上的近亲繁殖,基因已高度纯合化。纯合子的纯度在理论上接近最高点,基因已有98.6%以上完全纯合,仅有1%左右不纯合,因此近交品系动物的基因是一致的,遗传组成亦相同。在一个近交品系内所有动物的各个基因位点都应该是纯合子,这样的个体与该品系中任何一个动物交配所产生的后代也应该是纯合子,在这些动物中没有暗藏的隐性基因。
(二)遗传稳定性。由于近亲繁殖增加了在特定部位纯合子互相配合的可能性,因而减少了遗传变异,基因型可长期处于稳定状态,这种相对稳定性来自纯合性。因为基因高度纯合,所以纯合子基因可以极稳定地传给后代。如DBA系已维持好60多年,C57BL系已维持了50多年,但至今仍与原品系极相似。当然近交系动物有时会因突变造成基因改变而发生变异,是为例外。因此,近交系动物在遗传上是相当稳定的,遗传上的变异仅发生在少量残留杂合基因或基因突变上,而这种机率非常低。如果品系在被确认为近交系后坚持近交,同时辅以遗传监测,有时地发现和清除遗传变异的动物,保持近交系动物遗传稳定性是绝对没有问题的。
(三)同基因性。基因型或称遗传型是一切遗传基础的总和,是内在的遗传本质。同基因性,是指一个近交品系中所有个体在遗传上是同源的。由于基因高度纯合和基因型相当稳定,而致个体间极为相似,即同一品系内具有基本相同的遗传组成和基因点,也就是基因型相同。这种遗传上的均质性可采用组织移植方法来检测,也可用一只动物检测群体的基因型;从子代群体中检定母系群体。
(四)表现型的均一性。表现型是基因在环境因素作用下表现出来的、可被直接观察到的亲代的性状。在相同环境因素的作用下,由于遗传是均质的,所以其表现型是均一的,反应性是一致的。近交品系动物的性状如肿瘤发病率、形态学特点、血型和组织型、对药物的反应、甚至行为的类型等都可高度遗传,均衡一致远比远交系为强,因此可用较少量的近交品系动物,达到统计需要的精密度。
(五)个体性。由于不同品系都具有不同的遗传组成和生物学特性,所以不同品系各有不同的特性。目前国际上公认的250个近交品系小鼠间,均各有明显的区别,有其不同的反应性和敏感性,如自家免疫病鼠、白内障鼠、脑积水鼠、多尿症鼠、缺乏免疫球蛋白M鼠、对胰岛素敏感鼠等等。因此,选择近交品系鼠时,必须注意其是否适合各项研究设计的需要,决不能认为近交系动物遗传均一、基因型相同、反应性一致,就随便选一个品系来做实验。
个体性从整个近交系动物来看,每个品系在遗传上都是独特的,这表明在相当广泛的特性上,有些品系可能自发一些疾病,成为研究人类疾病理想的模型。在某些情况下,品系间的送别显示在量上,而不是在质上,而这一点在研究上也非常有用。因为以此可在很多的近交系中筛选出对某些因子敏感和非敏感的品系以达到不同的试验目的。
(六)分布的广泛性。许多近交系在国际上广泛分布,从而有可能在世界各国之间进行比较研究。这在理论上意味着不同地区、不同国家的科学家有可能去重复或验证已取得的理论和数据。但因环境变化可引起遗传变异,因此,环境条件应力求尽可能一致。如同一近交品系的两只动物,一只吃完善的饲料,另一只吃营养不足的饲料,则二者可发生不同的变化。
(七)资料可查性。近交品系的一个最有价值的特点是最常用的品系都具有相当数量的背景资料,由于近交品系动物在培育和保种的过程中都有详细的记录,加之这些动物分布广泛,经常使用,目前已有相当数量的文献记载着各个品系的生物学特征,这些有关品系的特征、寿命和自发性疾病等资料,对研究工作选择品系是极为重要的,而且这些基本数据对于设计新的实验和解释所得结果提供了便利条件。
(八)可分辨性。每个近交品系都具有自己独特的生化标志基因等特点,研究工作者可定期进行检测,以识别所使用的近交品系动物是否可靠。方法有:生化位点法、皮肤移植法、毛色基因法和下颌骨测量法。由于绝大多数近交品系在很多遗传位点上已有了分型,如果掌握了这些方法,可以根据这些位点的分型轻而易举的将混合在一起的两个外貌近似的品系分辨出来。
二、使用近交系的优点
使用实验动物进行各项科学研究的一个关键问题,就是怎样使动物实验的结果正确、可靠、有规律性、重复性好,从而精确判定实验结果、得出正确的结论。使用一般普通饲养的实验动物是采用任意交配的繁殖方法,所产生的动物个体差异较大,所以必然影响实验结果的均一性,有时难以判定实验结果。选用近交系动物作实验就能克服这些缺点,满足实验研究的需要。采用纯系动物作实验,具有下列主要优点:
1.具有相同的基因型,表现型也一致,所以其反应是一致的,实验结果正确、可靠。由于连续近交繁殖,同一近交系的各个体具有相同的基因型,在相同的环境条件下又具有相同的表现型,故其性状即其各种生物学特性比较一致,对外来刺激反应也一致。
2.各品系均有其独特的特性,根据实验目的可选用不同品系来作实验,实验重复性好,所用动物少,实验周期短,节省人力、物力和时间。
3.国际上分布广泛,不同国家的科研单位由于使用同一近交系动物所取得的结果是相似的,便于国内和国际间学术交流和实验重复。
4.可以作为有价值的病理学模型,如有致癌品系、抗癌品系、致白血病品系,嗜酒性品系、易抽搐品系等,是研究人类疾病的重要实验材料。
5.它是标准的实验材料,动物生长发育到一定时间就有一定的规格。
6.有大量的历史资料可查,每个品系均有其详细的遗传学资料,遗传背景明确,其生物学特性、生理生化特点、常见疾病(包括自发性疾病)等都有过系统的研究,便于研究者查阅和选择应用。
三、近交系动物在生物医学研究中的应用
近交系动物首先应用于遗传学研究,以后又为肿瘤和免疫学家所重视。在肿瘤的研究工作中应用最广泛,培育的品系也最多,对肿瘤的病因学、发病学、实验治疗和新抗癌药物的研究等都发挥了重要作用。所以近交系动物的建立已受到世界各国医学科学研究工作者的重视。正如化学家需要分析纯的化学药品、物理学家需要高度精密的仪器一样,近交系动物对医学研究是至关重要的。伦敦大学古勃宁说:“……生物学中近交的应用和化学中应用分析平秤的意义一样重要。”波尔法氏说“……癌的免疫治疗的最好途径的发展与1952年开始和发展起来的近交系动物是有关的”。
随着医学科学研究的飞速发展,近交系动物的培育及应用愈来愈被人们所重视,为适合不同课题研究需要而培育的近交系动物品系也愈来愈多,在医学、生物学、药物学等各个领域内的应用也日益广泛。生物制品、药品、食品等产品检定,疾病诊断、生理、病理、肿瘤、免疫、内分泌等学科的研究工作中都需要使用近交系动物进行各种实验。
近交系小鼠目前在国内许多科研单位已得到重视和应用。尤其在遗传、肿瘤、免疫、放射、白血病等研究中应用较多,其中在肿瘤、白血病研究中的应用更多,可以挑选致癌系小鼠进行致癌,可以在同一时间内获得许多生长同肿瘤的动物,然后进行各种种瘤的理论和防治研究。
在生物医学实验研究中应用最广泛的是近交系小鼠,它可以根据各种特殊实验的需要培育各种品系,如致癌鼠、抗癌鼠、糖尿病鼠、白血病鼠、先天性肌肉萎缩症鼠等。近交系小鼠应用也较多,近交系的金黄地鼠、豚鼠以及兔狗等也有应用。1975年出版的第三版《国际实验动物索引》中公布的的交系动物品系数,小鼠388系,大鼠130系,豚鼠8系,家兔2系,黑线仓鼠2系、金黄地鼠38系。现在世界上已经有纯品系小鼠500多个品系,大鼠包括亚系在内约200多个,豚鼠12个,家兔6个。
从1952年以来,国际近交系小鼠标准命名委员会(Committee on Stanardized Nomenclature for Mice)对承认的近交系小鼠进行命名,每隔4年在美国肿瘤研究杂志上公布一次,先后于1952、1960、1964、1968、1972、1976、1980和1984年共公布过八次。第一次1952年公布了124个品系,第五次1972年已发展到244个品系,1976年为252年品系,1980年为250个品系,1984年为250个品系,我国培育的615、津白Ⅰ和津白Ⅱ小鼠也得到承认并被列入。
近交系动物的广泛应用,极大地提高了实验结果的正确性和重复性。以近交系小鼠为例,目前已广泛应用于的肿瘤研究课题(见表8-1),也广泛应用于基础和临床医学的各类实验研究(见表8-2)。
杂交瘤(Hybnridoma)合成单克隆抗体(Monoclonal antibodies)是近年来生物医学中一项重大的突破,将对人类肿瘤的治疗,传染病的防治和诊断及免疫机制的研究等方面产生巨大的影响和变革。这项新技术选用的主要实验材料就是BALB/C近交系小鼠,单克隆抗体研究就是从BALB/C小鼠骨髓瘤开始的,近几年来单克隆抗体的研究,又有了新的发展。双特异性单克隆抗体(bsMCA)的研究,是英国剑桥大学分子生物学实验室Milstein博士正在进行的研究(Milstein是单克隆抗体技术创始人之一),这在国际上刚开始,国内尚未展开此项研究。制备又特异性单克隆抗体就必须要用Lou株近交系大鼠,因为Lou株近交系大鼠制备单克隆抗体时,其腹水量比用BALB/C小鼠大几十倍,能较好地解决单克隆抗体的大量制备。双特异性单克隆抗体技术和杂交~杂瘤技术是单克隆抗体技术的新进展,bsMCA可代替交联剂和酶标技术,在免疫组化和免疫测定技术上有广泛应用,在癌症的导向治疗和体外免疫扫描诊断上也有广阔的应用前景。
第三节 近交系的培育技术
一、培育目标
培育目标分定向、不定向两种,定向是予先确定好要获得什么样的品种,然后有计划地进行培育,这种方法能在较短的时间里获得优良的品种。定向时有以细菌感受性为选育指标,如小鼠对沙门氏菌的感受性;也有采用遗传指标,如性成熟、初产日令、妊娠间隔、产仔数、哺乳量及离乳时体重等。不定向的是无意识的选择,预先交没有想要培育具有哪些特点的新品种,只是人们自然把比较有价值的留下,让它繁殖,去掉价值不大的品种,这样无意中起了选择的作用,这种选择过程是比较缓慢的,待纯化后回过头来做实验,测定其特性。
二、种鼠的选择
(一)基础种群的选择
1.从野生鼠中选择。可能具有实验小鼠所没有的未知遗传特性,但由于野生鼠需要经过一段驯化时期,有时人工繁殖有困难,所有这种情况应加以考虑。
2.从起源于没有经过的新繁殖的近交群中选择。由于其个体之间存在着某种程度的遗传变异近亲繁殖和选择,把可能获得并需要的特性固定下来。
3.从杂交群中选择。这种杂交群包括,近交系与近交系之间;近交系与远交系之间;远交系与远交系之间;野生鼠与远交群之间的各种类型的杂交。尤其是“三元”即三个不同品系之间,“四元”即四个品系之间的杂交,有可能通过交配而形成多种的基因组合,从中选出具有一定特性的鼠种作为基础群。
4.如果其它实验室和保种单位有质量较好基础种群种子,并有相应的检测数据证实其确为好种子,也可直接引起,这样较为简便易行。但引进时必须了解和掌握必要的资料,如家谱号(Pedigreed No.)及其历史来源、交配方式、近交代数、遗传组成、生物学特性等。
(二)初代种鼠的选择
在选好基础 种鼠以后,进一步选出初代种群是极其重要的。
1.首先检查个体形态,如毛色、体型以及对外界刺激反应的敏感性等,有目的的选择后,即可进行繁殖。
2.对初代种群所产生的仔代,应进行病原微生物和药物感受性以及肿瘤诱发率等方面的检查,必须选留那些经过检查,而且有一定特性的双亲所生下的1~3胎仔鼠,作为下一代生产的种鼠。
3.第二代以后种群的选择。与选初代种群基本相同,但应注意二点:
(1)除注意其所需特性以外,还应注意其初产年龄、产仔数量、哺乳能力等方面的繁殖特性,应选择质量优良者。
(2)尽量在5~7代中选择具有所需特性的种鼠进行交配,因为5~7代是兄妹交配选择效果最好的代数,因此5~7代交配的组合尽可能大些,超过10代选择效果则较差。
三、培育方法
培育近品系的重要手段,一是交配繁殖,二是人工选择,两者经常是同时进行的。交配繁殖的目的在于使存在于基代双亲(即用于培育近交系的最初一对小鼠)中的不同遗传位点的不同基因通过连续许多代的重新组合达到纯合的程度,并保持足够数量的后裔可供交配时选择。人工选择的目的是挑选符合培养需要(有繁殖能力或其它的特殊性状)和淘汰不需要的个体。
(一)近交的交配方式
开始培育近交系种群和引起的近交系原种的维持,一般采用以下三种方法:
1.单线法:从近交系原种选出3~5个兄妹对进行兄妹交配,从中选出生产能力最好的一对进行繁殖,然后从中选择一对作为下一代生产的双亲,如此一代代的延续下去。这个方法生产的个体比较均一,但选择的范围太小,由于只有单线的仔代,易发生断代危险。
2.平行法:选3~5对兄妹对,每个兄妹对都选留下一代种鼠,一代代的延续下去,这个方法生产的个体不太均一,但选择范围大,易发生分化。
3.选优法:这个方法保留了上述两个方法的优点克服了两个方法中的缺点,是个较好的保种方法。假定每代选6对,每对都选自同一双亲的仔代同胞兄妹,在繁殖过程中,每一代均保持6对,当某对出现不孕或生产能力降低,而不适于继续繁殖时,则可从另一对所生的后代中选择优良者加以代替。近交系原种动物的维持方法见图3-2。
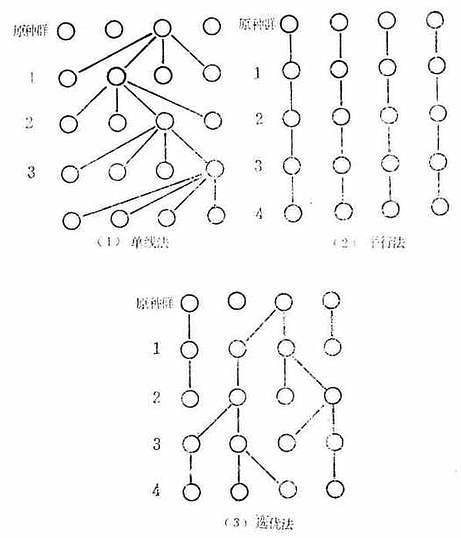
图3-2近交系原种群(基础群)的维持方法
(二)交配繁殖
1.近交系的培育要在专用固定的饲养室内进行。为了避免别的鼠系的进入和防止可能发生的感染,应有严密的隔离设备和措施。如墙壁门窗都不能有缝道,饲养罐盖子要十分严密,不能让任何小鼠随便进入。外观相似种系不同的动物应分室饲养。工作人员进入饲养室时需带口罩,非饲养人员不能随便进入。饲养罐、饮器、食具等使用前都要灭菌消毒等。
2.应该严格遵照单对配对,父母同罐的要求。这样便于随时检查和发现问题。而且雄鼠不离开雌鼠可以减少搞错谱系的危险,雄鼠也不干扰幼鼠的哺育和成长。一般不应采取一雄二雌的交配饲养方式,因为两个雌鼠可能同时怀孕,有时难于辨认年龄相仿的仔鼠与母鼠的关系。
3.对同系的每个鼠都应有详细明确的谱系和户籍记录:包括饲养罐号,同对的雌雄鼠号码(可根据父母鼠号码、胎数、同胎中第几号等确定)、培育代数、生育日期、胎数、产雌雄仔鼠数、断乳后存活的雌雄仔鼠数,选作传代交配的仔鼠号等等。记录不全或户籍登记错误往往会造成无法挽救的损失,特别是当这个错误未被及时发觉时。如果对一个动物谱系身份有怀疑,应立即予以丢弃。另外,所有饲养罐也要加贴有明确记录的标签并随时进行检查,发现任何情况都就立即记入户籍记录。
4.选用第一、二胎同胎仔鼠作b×s交配比b×o交配更易处理,而且常常繁殖的更好。如有必要也可用同父母不同胎的仔鼠配对。譬如在一胎仔鼠的性别完全或大部分相同的时候,经过很多代的近交繁殖以后,有时也可以用亲缘关系较远的堂(表)兄妹进行交配。当然这时近交系数要退回到它们的共同祖先(对堂兄妹交配来说是退回到祖代)这一代的F值。但如这种做法只是为了想提高繁殖较差的生育率,那么倒不如停止这条线的繁殖,另找一个生育较高的亚系作为近交繁殖的新起点更为妥善,因为前面的做法即使能够培育出一个近交系来,它的后裔也往往难以提供足够的实验动物数量。
5.在近交系培育过程中,为了保证对生育力有一定的选择范围(生育力的选择常常需要挑选多产和断乳后存活的仔鼠较多的亚系进行交配繁殖),每一代都应保留一定数目(3~4个)的亚系,以后随着代数的增加,除留下最后2~3代分出的亚系外,其余可陆续淘汰或抛弃,直到最后留下一个可追溯到基代的品系。因此除了记录下谱系和户籍之外,常常还需要随时绘制象下面的那些谱系图,如图3-3,以便分清或随时终止某些亚系。
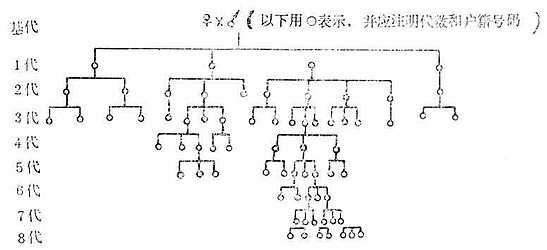
图3-3 近交系培育过程中动物的谱系图
提供交配繁殖的亚系选择应特别注意:
(1)这个亚系应具有培育所需要的性状。
(2)至少与准备终断的亚系有差不多的生育率和生活力。在繁殖良好的种系中一般在第3~4代以后就可终断其它的亚系而只留一个亚系。如果子裔不太多,至少需传5~6代甚至10代以后才能这样作。
6.在完成20代b×s交配繁殖而建成一个近交系之后,要繁殖足够的动物提供实验需要,同时又不放弃选择最适合的动物进行繁殖并尽可能的保持遗传的一致性,最好的作法是从近交系中取出几对较好的动物继续照上面的办法进行b×s交配传代,并保持谱系和户籍记录,同时把其余的动物另室饲养,让最后三代任意交配,加速繁殖。可以每罐放1雄4雌,待受孕后取出母鼠,用它们的后代提供实验需要。开始时,应注意多保留雌鼠。
第四节 近交品系的命名法
近交品系的命名,国际上已有统一规定,由国际实验协会(ICLA)负责,自1952年以来,近交系小鼠标准命名委员会对承认的近品系小鼠每四年公布一次,1984年第八次公布了250个品系,其中包括各个系或株(stock)的历史、来源、独特的生物学特性等介绍,以及人名、单位名的规定写法和遗传背景、基因组成、缩写符号及其相应品系的标准命名等。这些资料供全世界参考,在进行新的命名时可避免重复及混乱。
近交系动物国际命名的规则是根据动物的来源、历史和培育经过而命名的,用一系列的字母及数字来表示。
一、近交系的命名法
1.一般均用一个或几个大写英文字母表示,如A、AE、BA、AKR、DBA、NZB、STAR、NMRT、NYLR、NZBR等。
2.有些品系可在大写英文字母间加入一些阿拉拍数字来表示,如C3H、C57BL、C57BR、C57L、CC57W、CC57BR等。
3.非正规的命名,如果已广泛为国际所共知者,则可保留沿用,如:“129”、“101”、“615”等。
4.近交代数表示,一般是在品系符号后括号里写上代数,并在代数前加写“F”(Filial的缩写),例如A(F78)。如果由于资料不全,可以写上已知代数,在代数前面加下问号,如AKR(F?+10)如系从其它实验室引入的近交系或亚系,又经自己实验室若干代的繁殖,其近交代数的表示方法是在F符号后,先标明引入时的子代数,再加上自己实验室繁殖的代数,如C57BL/Jnga(F73+26),即表示此亚系是73代时引入的,又经自己培育了26代。
5.书写品系命名符号要写全,应使用公布的全称,不能随便缩写(这里指的是写论文报告,在材料方法一栏里一定要把品系命名符号写全。这样才能说明所指的是哪个品系或亚系)。例如,C57这个缩写可意味着七、八个特征不同的品系(如C57BL及其亚系6个,C57BR/cdJ,C57L)。只写C57BL就包括有几个特征不同的亚系(如C57BL/1,C57BL/6J、C57BL/6N、C57BL/10J等)。所以,若只写C57或C57BL都是品系名称未书写完全。同样,只写CBA亦为不确切的书写法,因为它也有若干个特性不同的亚系,如CBA/J是有视网膜退变基因的,而CAB/ca则没有这种基因,两者对放射线的敏感性也不一样,组织相容性也不同,至少有5个位点不同的。
6.国际上公认的品系缩写:已公布的品系缩写有:AKR-AK,BALB/c-C,CBA-CB,C3H-C3,C57BL-B,C57BL/6-B6,C57BL/10-B10,C57BR-BR,C57L-L,DBA/1-D1,DBA/2-D2,HRS/J-HR,RⅢ-R3。
7.品系符号的优先权:任何品系或亚系命名均应参照国际命名委员会的规则命名,如果出现两品系命名符号相同,则保留先正式公布者,并以发表《Mouse News Letter》(小鼠新闻通讯)或《Inbred Strani of Mice》(近交系小鼠)两个刊物上的品系灵为准。但一些著名的、国际广泛使用的品系则不在此限。
二、亚系的命名法
(一)亚系的定义和形成
亚系(Substrain)是由同一个近交系分离出来的具有各不相同特性的品系。亚系可由下列途径形成:
1.在同一研究室里,从近交品系中分离出来,从兄妹交配繁殖达8~19代之后分开饲养,分开后不与其它品系混交,再继续兄妹或亲子近交12代以上者。如此可获得具有一定特殊性状的品系。
2.一个近交品系从一个研究机构或研究者转送到另一个机构或研究者哪里,经相当代数饲养,并与其它品系混交者。
3.当发现有某些基因变异时,可培育成具有某些特殊性状的亚系动物。
(二)亚系的命名表示方法
1.在原亲本品系名称的后面加一条斜线“/”,再在斜线下标上适当的亚系符号,亚系符号可分为两类。
2.一类亚系符号是用研究人员或研究机构名称的缩写英文字母,第一个字母须大写,随后字母要小写。
(1)用研究人员名称的缩写字母表示,如A/He,这里,He即做为亚系符号,He是人名Heston的缩写,即美国国立肿瘤研究所W·E·Heston博士,A/He可称为A系的Heston亚系。
(2)用研究机构名称的缩写字母表示,如A/jax或J,Jax或J为美国著名的肿瘤研究机构Jakson实验室。
(3)老亚系中出现新亚系,即当一个亚系从一地方迁移到另一个地方,即被另外一个人培育而成新的亚系时,最好老名也保留,可在老亚系符号后面加一新亚系符号。如YBR/HeW;就是将YBR小鼠的Heston亚系,迁移到Wilson处而成的一个新亚系。这样就能通过命名表示系发展的历史和变迁顺序。如果把前一节删去,写成YBR/Wi也可以,但就割断了历史,应在记录中加以说明。又如,C3H是小鼠高癌品系,Strong氏发现的第一亚种是40年前发现的,目前已繁殖了50或更多的世代,现在世界上至少已有四个大的亚系;①最早的亚系是Strogn发现的,故以C3H/ST表示;②以后,1930年又由Anderant氏分离出,故叫C3H/An;③1931年Bittner氏引入后培育到13代以上,又叫C3H/B;④1941年又由Heston分离,故又定名为C3H/AnHe或叫C3H/He。现已证实这些亚系之间虽然都是由C3H来的,但已发生了重要的变异,有人实验指出C3H亚系间对于病毒诱发白血病的敏感性差异明显,C3H/B有18.4%产生白血病,而C3H/He小鼠只有1.2%产生白血病。
3.另一类亚系符号是用阿拉拍数或小写英文字母作为亚系的符号。
(1)用阿拉拍数字表示:一般用于一个品系或亚系分离形成两个亚系或两个新亚系,并仍在一个实验室饲养,可在原品系符号末尾用数字作为亚系符号,其前可加(也可不加)斜线,但一般都加斜线,如A/HeCrgl/1及A/HeCrgl/2,这是美国加利福尼亚大学癌肿研究遗传实验室在同一研究室里形成的两个亚系,以1和2为新亚系的区分,当然也可写成A/Crgl/1和A/Crgl/2。
(2)用小写英文字母表示:当一个品系(或亚系),分离成两个亚系(或新系)而远未达到基本近交繁殖以前,其符号可用小写英文字母表示。如C57BR/a和C57BR/cd,这两个亚系分别是在全同胞交配到第9代和第13代分离时而命名的,如果一个品系(或亚系)分离形成的两个亚系(或两个新亚系),兄妹交配达20代以上者,就不要用数字或小写字母来命名。纵然有少数例子,如DBA/1和DBA/2,因其在国际上已被广泛沿用,算是例外。
4.因代乳、卵或卵巢移植所形成的亚系。
(1)代乳(Foster Nursing)的亚系以“f”作符号,写于原品系名之后。如C3Hf表示这C3H是通过代乳养大的亚系。还可写得更清楚,把寄奶雌鼠的品系名也写在f之后,如C3HfC57BL,或缩写成C3HfB,即表示C3H是由C57BL代乳的。
(2)受精卵转输(Dva Transfer)或卵巢移植(Ovary Transplant)用“e”或“o”(即卵eggs或Ova第一个字母)符号表示。如AeB,即A的受精卵转输到C57BL母鼠的子宫。另外还可以与其它亚系符号联用,如A/HefB。
当两个亚系之间连接有代乳号“f”或转输符号“e”或“o”时,为了避免符号混乱,可以将f或e或o用希腊书法写在两个品系或亚系符号之间较下方。例如,A/HefB,即A的Heston亚系,由C57BL代乳。
5.复合亚系符号(Compound substrain symbols)
由于代乳或导入一个外来基因而形成的复合株则需用复合亚系符号。这种符号要体现出品系发展的历史和时间顺序。譬如采用代乳、卵或卵巢移植、基因突变、导入外来基因以及是谁培育或是谁保种等等都可以通过复合亚系符号按历史时间的顺序表达出来。
(1)亚系符号写在品系符号后面,例如BALB/cf,DBA/2eB,C3H/Ha-p。其中C、2、Ha为亚系符号,亚系符号后面的符号f、e、p分别说明代乳,受精卵转输各突变种。
(2)亚系符号后面记上处理过程或导入基因的符号,在亚系符号后面的位置上涉及代乳或受精卵转输以及目前保种者或研究室名称的符号。例如DBA/2eB/De或DBA/2eBDe,即这个亚系是由Deringer把DBA/2的受精卵转输C57BL,由Deringer保种。又如C3H/Hef/Ha,即C3H/He由Heston用代乳鼠代乳,目前由Hauschka保种。又例如CBA/Ca-se/Gn,即Carter的CBA亚系具有Se基因突变(短耳Shortear),由Green保种。
这里要说明的是,类似DBA/2eBDe,在“De”这个位置上的符号,其含意有三;代表培育这个品系的人,保种者,或兼而有之。即究竟是由Deringer建立此品系,还是由它保种或二者兼者,这需要由记录说明,所以应当登记清楚。
6.冰冻保体传代的近交系小鼠命名规则。
(1)凡冰冻保存的品系,即在品系标准命名符号之后加小楷英文字母“p”,即Preservation by freezing(冰冻保存)。
(2)若冻存胚胎移植到同一品系母鼠子宫内代育时则命名符号不需改变。例如:C57BL/6p 即指C57BL/6的冻存卵移植到C57BL/6母鼠子宫内代育。
(3)如果冻存胚胎移植到不同品系母鼠的子宫时,应在被代育品系标准命名符号后面加上e ,例如:C57BL/6pe、CBA/H是指C57BL/6的冻存受精卵移植到CBA/H母鼠的子宫里(其中的H是指Har Weu亚系的符号)。
(4)亚系符号中应表示出冻存的人或实验室符号,其人名、单位名可接着写,如C57BL/6peCAB/HBy,其By即表示此品系是由D·W·Bailey冰冻的。
(5)近交代数、冰冻保存与解冻年份加在p后面的括号内,冰冻前后代数写在括号外左右两侧。如F37+(p1975-1977)+F16,是指1975年近交繁殖达37代时作冰冻保存,到1977年解冻,自解冻后又进行近交繁殖达16代。
如果冰冻与解冻是由不同的人或单位所作,则可以在括号里说明,如(p175By-1977H),是指1975年由D·W·Bailey冻存,到1977年由Harwell,(是英国M·R·C放射生物学部门所在地)解冻。
7.近交品系动物综合表示方法
DBA/1fLACA/LAC
DBA表示小鼠近交品系的名称;
“1”表示亚系;
f表示代乳;
ACA表示代乳母鼠的品系名称;
LAC表示培育单位代号。
上述一串符号初看起来很复杂,但只要按上述介绍的命名规则细加分析,就很容易看明白,这是由英国实验动物中心(LAC)用自己培育的LACA小鼠代乳而培育的DBA/1小鼠。有这样的命名法,大家就可以清楚地知道这些的交系动物的来源、历史和培育经过。
又如:C57BL/6J
“C”代表Cold Spring Harbor Labaratory;
“57”代表第57号雌鼠;
“BL”代表BLACK(黑色);
“6”代表亚系的数字;
“J”代表Jackson Laboratory。
8.亚系常用的代号。
An:美国国立肿瘤研究所的H·B·Andervont博士。
BCr:Birmingham大学肿瘤研究所。
Cd:比利时Libre大学Albert Claude博士。
COBS:Caesarean Originated Bustained Animals,用无菌手段取出后养在封闭系统。
Crgl:美国伯克莱加利福尼亚州大学肿瘤研究所遗传实验室。
CRJ:日本Charles River Japan,Inc公司。
FO:美国Michigan州Detroit大学生物系P·Forsthoefel博士。
Gr:英国伦敦大学动物系H·Grunebery博士。
Gro:荷兰Groninge大学遗传研究所。
H:英国医学研究协会放射医学部门所在地Harwell。
Han:德国Hannoverlinden Versuchstierzueht中心研究所。
He:美国国立肿瘤研究所的W·E·Heston博士。
H-2:Histocompatibility-2allele组织相容性抗血清Ⅱ型的特性。
ICIA:International Committee on Laboratory Animals Science。国际实验动物科学委员会。
ICR:美国国立肿瘤研究所(Institute for Cancer Research)。
ICRC:印度癌肿研究中心(India Cancer Research Centre)。
J或Jax:美国杰克逊实验室(Jackson Memoriel Laboratory)。
JCL:日本柯力亚(Clea)公司(株式会社)。
JCR:日本国立肿瘤研究所。
Jic:日本(财)实验动物中央研究所。
JMC:日本东京大学医科研究所(田鸠加雄)。
Ka:美国加利福尼亚Stanford大学医学院放射原系H.S.Kaplan博士。
Ki:美国Ohio大学医学院Kirschbaum纪念研究室。
Lac:英国M.R.C实验动物中心(Laboratory Animals Centre.Carshalton,Surrey,England)。
Mcl:英国苏格兰爱丁堡动物遗传研究所A.Mclaren博士。
M.K:日本北海道大学理学部(牧野佐二郎)。
MS:日本(Misima)国立遗传学研究所(吉田俊秀)
N或NIH:美国国立卫生研究院(National Institute of Health。)
Pi:美国Utan大学医学院的H.I.Pilgrim博士。
Rd或Rol:法国巴黎G.Rudali博士。
Scr:美国Lajolla Calif Scripps临床和基础研究基金会实验病理系。
S.P.F:无特殊病原体动物或称无致病病原体动物(Specific Pathogen Free,Animals)。
Umc:美国Minnesota大学医学院生理系。
W.H.O世界卫生组织(World Health Organization)
Y:苏联Yurlov邮政实验动物研究所遗传系。
第五节 近交系的新进展
一、重组近交系
重组近交系(Recombinant Inbrad Strain)的发展和使用,是哺乳类动物遗传学中的最重要的发展。这是近十年发展起来的,是以两个无关的高度近交品系进行交配,产生F2代后,再行全同胞交配达20代以上而育成的一个近交系列组动物。该品系动物既具有其双亲品系的特性,又具有重组后一组内和每个重组近交系的特征,因此已广泛应用于新的多态形基因位点和新的组织兼容性位点的鉴定、多态形位点的多效性和多态形位点的连锁关系的研究和探测,以及临界特性的遗传分析,除此之外,也用于寿命、自发性和诱发性疾病感受性的研究,还用于生理学、药理学、形态学和行为特性的生物统计分析等方面的研究。
重组近交系名称的书写方法是在两个亲本品系名称之间加一个“X”符号(居中而不留空间)来表达,品系名称用缩写形式。如BALB/cByXC57BL/6By,记为CXB;C57BL/6JXDBA/2J,记为BXD;C57BL/6JXC3H/HeJ,记为BXH;C57BL/6JXSJL/J,记为BXJ。对于其一组内的不同品系应用阿拉拍数字加一对开线“-”加以区别。如BXD-5,BXD-30;BXH-19,BXH-2;BXJ-1,BXI-2等。早年以大写英文字母来区分,如由两个无关的、近亲程度较高的BALB/cBy(缩写为C)和C57BL/6By(缩写为B6)品系之间的互交,经过20代以上的兄妹交配而育成的,并命名为CXBD、CXBE、CXBG、CXBH、CXBI、CXBJ和CXBK。最近均已不用大写英文字母而用数字表示,如AKXC-1,就是AKR和BALB/C品系互交后,经兄妹交配而育成的重组近交系之一。已培养成功和正在培育的重组近交系可见表3-3。
表3-3 已培育成功的和正在培育的重组近交系
| 亲本 品 系 |
重组近交系命名 |
品系 数 |
参 考 资 料 |
| BALB/cBY,C57BL/6By |
CXBD到CXBK |
7 |
Bailey(1971) |
| AKR/J,C57L/J |
AKXL-1到AKXL-38 |
21 |
Taylor和Mcier(1976) |
| SWR/J,C57L/J |
SWXL-到? |
8 |
Taylor(1976) |
| C57BL/6J,DBA/2J |
BXD-1到? |
24 |
Taylor(1976) |
|
|
|
Taylor et al(1973) |
| C57BL/6J,C3H/HeJ |
BXH-1到? |
14 |
Taylor(1976) |
根据1981年Bailey的统计资料看,目前至少有28个重组的近交系列组,见表3-4。
表3-4 RI品系的来源与建立(Bailey,1981)
| 亲 代 品 系 |
| 序号 |
组合品系 |
雌 性 |
雄 性 |
动物数 |
繁殖代数 |
来 源 |
| 1 |
AKXD |
AKR/J |
DBA/2 |
30 |
11-14 |
Heiniger,Taylor(Jax) |
| 2 |
AKXL |
AKR/F |
C57BL/J |
18 |
15-40 |
Taylor(Jax) |
| 3 |
AXB |
A/J |
C57BL/6J |
10 |
7-16 |
Nesbitt(UCSD) |
| 4 |
BNXAKN |
C57BL/6N |
AKR/N |
12 |
11-25 |
Nebert(NIH) |
| 5 |
BNXC3H |
C57B57/6N |
CeH/HeN |
12 |
12-17 |
Nebert(NIH) |
| 6 |
BRX58N |
C57Br/cdJ |
B10,D2(58N)/Sn |
11 |
20-27 |
Taylor(Jax) |
| 7 |
BXA |
C57BL/6J |
A/J |
11 |
7-17 |
Nesbitt(USCD) |
| 8 |
BXD |
C57BL/6J |
DBA/2J |
24 |
29-44 |
Taylor(Jax) |
| 9 |
BXH |
C57BL/6J |
C3H/He |
13 |
33-41 |
Taylor(Jax) |
| 10 |
BXJ |
C57BL/6J |
SJL/J |
2 |
23-33 |
Taylor(Jax) |
| 11 |
BXLG |
C57BL/10J |
LG/J |
7 |
14 |
Haber(BNL) |
| 12 |
CXB |
BALB/CAnNBy |
C57BL/6NBy |
7 |
60-70 |
Bailey(Jax) |
| 13 |
CXD |
BALB/cJPas |
DBA/2Jpas |
10 |
13-21 |
Guenet(pasteur) |
| 14 |
CXJ |
BALB/cst |
SJL/J |
12 |
10 |
Guckeler(salk) |
| 15 |
CX8 |
BALB/cwt |
C58/J |
9 |
4-7 |
Stevens(Jax) |
| 16 |
LTXB |
LT/sv |
C57BL/6 |
4 |
16-21 |
Stevens(Jax) |
| 17 |
LXB |
C57L/J |
C57BL/6J |
3 |
23-33 |
Taylor(Jax) |
| 18 |
LXHR |
C57L/J |
HRS/J |
17 |
10-18 |
Stoner(BNL) |
| 19 |
LXPL |
C57L/J |
PL/J |
5 |
12-16 |
Taylor(Jax) |
| 20 |
NXC |
NZB/NBom |
BALB/cJ |
28 |
10-12 |
Krog(Fibiger) |
| 21 |
NXSM |
NZB/BINJ |
SM/J-ala |
18 |
15-22 |
Eicher(Jax) |
| 22 |
NX129 |
NZB/BINJ |
129/J |
8 |
10-15 |
Taylor(Jax) |
| 23 |
NX8 |
NZB/lcr |
C58/J |
13 |
18-24 |
Riblet,Weigert,Johnson(Icr) |
| 24 |
SWXL |
SWR/J |
C57/J |
7 |
18-40 |
Tayor(Jax) |
| 25 |
SWXN |
SWR/J |
NZB/BINJ |
12 |
6-8 |
Datta(Tuflts) |
| 26 |
9XA |
129/Sv-ST |
A/HeJ |
13 |
6-7 |
Stevens(Jax) |
| 27 |
58NXL |
C57L/J |
B10.D2(58N)/Sn |
5 |
17-23 |
Taylor(Jax) |
| 28 |
129XB |
129/sv |
C57BL/6JPas |
15 |
14-19 |
Cuenet(pasteur) |
二、同源导入近交系
将一个基因导入到一个近交系(通过多次回交)而培育成的新的近交系称为同源导入近交系(Congenic Inbred Strain),也称为“近交同类系”或“同源抵抗系”,简称IR系。这是Snell(1948年)的大发明,通过人工培育成功了IR系。例如,他用易感受与好发某肿瘤的近交系小鼠,导入另一品系对该肿瘤无易感性的基因(不同的组织兼容性基因),先杂交一次导入,尔后通过一系列的子代互交、测验以及回交的办法而育成IR。具体方法参看图3-4。图中的A系为易感受与好发某种肿瘤,在A系内移植A系肿瘤都能增殖生长,因为它们是同质动物,不起排异反应。如果将A系肿瘤移植到B系动物,肿瘤就不能生长,因为B系与A系之间的组织相容性基因不一样,所以B对A的肿瘤起排异反应。从A品系的角度培育对A系肿瘤无易感性的IR品系的方法是,让基代A与B交配,生下子1代,再让子1代互交得2代。此时将A系肿瘤移植给子2代,选肿瘤不长的小鼠留种与A品系回交得子3代,再让子3代互交得子4代,此时再移植A系肿瘤给子4代。如此反复,一直进行到第12~14代,选不长肿瘤的小鼠以全同胞兄妹交配形式建立了A·B品系。此A·B系即为A系的IR系,两者也就是互为同类系。新培育的A·B系除组织相容性基因与A系同外,其它基因则基本相同,这样配对的品系可开展许多科研,且得到了重大的成果。按这个模式,可以培育出各种各样的同类系。
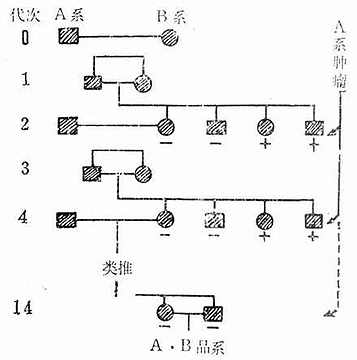
图3-4 Snell培育IR系小鼠的方法
三、异单基因近交系
相同近交品系的动物内,如果发生了单个基因的突变,而培育成的新的近交品系称异单基因近交系(Colsognic Inbred Strain)。譬如近交系小鼠129的种群中,某些个体发生了“肌萎缩症”的隐性遗传突变,由dy基因控制这个性状。这个突变育成了dy的突变品系,称为129-dy。这样129与129-dy便互交成为了同类系,因为它们两系除dy一对等位基因之外,其他基因都基本是一致的。这一对同类系是天然发生而通过人工选择育种的成功的,用这一对同类系来对肌萎缩症进行对比研究有重大价值。其他诸如在贫血方面的研究等等,也有许多类的例子。
同源导入近交系和异单基因近系通称为同类系,并称同源株(Congenic stock),是在一个近交品系内发生了一个重要的单个基因突变,或者通过一系列的多次杂交和回交把一个基因导入一个近交背景品系内部形成与原株相对应的同源株。
其命名是在原来品系或亚系名称符号之后加一连接号“-”,再写一斜体字的基因符号。如果这个基因是隐性的则字母全用小写,如果是显性的则第一个字母用大写,如DBA/Ha-D,129-dy等。
如果突变基因或导入基因仍处于杂合状态,则在“-”后添一加号“+”,再加基因符号,如A/Fa—+C,C3H/N-+Wj。
当同类系是采取中制杂合近亲繁殖的,则其分离位点是否写出随意,例如129或129-CcbC;SEAC-a+/+se或SEAC/Gn。
通过反复杂交将显性基因导入一个近交品系里,至少回交7代才能建立同类系,因此要在括号内标明回交代数,如C57BL/6-Wv+(N8)。(Wv是“活显性白斑”基因,是W的等位基因)。至于代数计数,是把第一次杂交(bybrid)的F1计算为第一代,第一次回交所产生的下代计算为第二代,以此类推。
同源株如在两个品系杂交中形成,其命名符号,一般是由最初杂交的两个品系的命名符号所形成,例如,B1O,D2/OSnN(特征是缺少补体5),O代表Old(老),BIO·D2/OsnN(特征是有补体5和脑积水者较常见),n代表new(新)。
我国现已引进的突变同类系(异单基因近交系)有:C57BL/6J-Ob/Ob,为先天性肥胖型动物模型,血糖偏高。C57BL/KS-db/db,为Ⅰ型糖尿病模型。C57BL/6J-bg/bg,为NK细胞缺陷的模型。
四、遗传工程与近交系的发展
遗传工程是70年代兴起的一门崭新的科学技术,它使人类进入了定向控制生物遗传性状的新阶段。
所谓遗传工程也就是根据人们的意愿,采用工程建筑的手法,按照预先设计的方案,借助于实验室手段将一个生物体的遗传物质定向地转移到另一个生物体中去,使后者获得人类希望的性状,成为一个新的“物种”。这们就打破了传统的、必须经过两性杂交的育种方法,使原来在自然状态下根本不可能发生有性繁殖的两种生物的基因结合在一起了。
遗传工程的概念可分为广义的和狭义的两种。广义的遗传工程包括细胞工程、染色体工程和基因工程三类,使用的是细胞生物学方法和分子生物学方法,包括细胞融合、细胞拆合、染色体导入和基因或DNA分子的转移等。广义的遗传工程又称为微生物工程或细胞工程。狭义的遗体工程仅限于基因工程。
早期的遗传工程的研究工作大多数是在病毒、噬菌体和细菌中进行的。因为这些生物的遗传物质比较简单,容易搞清楚遗传物质与生物性状之间的关系。随着知识的积累,技术的提高,现在越来越多的遗传工程实验开始在高等哺乳类动物身上进行了。这尤其促进了近交系小鼠的发展。
(一)细胞工程与近交系小鼠的发展
1.嵌合体小鼠(Mouse Aggregation Chimeras)的育成。嵌合体小鼠技术是由Tarkowski和Mintz分别在华沙和费城发展的(见图3-5)。他们首先用两对不同品系的近交系小鼠在同一时刻进行纯交,为了使其后代便于区分,常选择毛色不同品种的近交系小鼠例如选用白毛的SJL小鼠与黑毛色的BL/10小鼠。当受精卵分裂为8分裂球时,分别将它们从各自母体的子宫上分离下来。然后用蛋白酶消化分裂球外面的明带,使分裂球“裸露”。并在37℃的条件下,将来自两个品种的两个分裂球彼此接触,任其粘成一个具有双倍体积的早期胚胎。将早期胚胎继续培养到具有128~256个细胞的胚囊。这时,不同毛色品系的细胞相互混杂发育在一起。然后,通过手术把胚囊移植到寄养母鼠的子宫内,让它继续发育,直至出生。新生小鼠长出毛后,其毛色表现为既不象父亲SJL品系的全白色,也不象母系BL/10品系的全黑色,而是表现出黑、白条或块状的毛色。这说明新生鼠的组织是由“黑色”细胞和“白色”细胞嵌合而成的,是一只嵌合体小鼠。
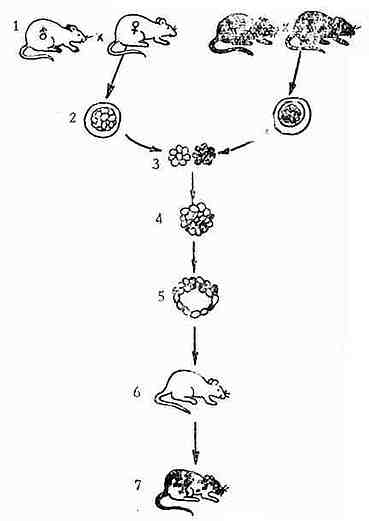
图3-5 嵌合小鼠育成
近交同类系动物也是近交系动物,它除了一小段带有可辩的目的基因染色体外,在遗传上与原来已建立起来的那个近交系完全一致。近交同类系动物的培育,是选用带有目的基因的个体与已经建立起来的近交系杂交建立起的。应用近交同类系动物可以研究多基因系统中一个基因的特殊作用,施耐尔博士(Snell)是第一个应用这种体系进行组织移植基因研究的,建立了同类抵抗系学说,为小鼠的主要组织相容性抗原的研究做出了巨大贡献,为此获得了1981年的诺贝尔奖金。
同类系动物的命名基于如下两种方式。一种是带有近交品系和供体品系的复合符号,中间带有一点,例:B6·AKR。第二种命名法是近交品系后面加上一横,然后标上供体品系特异的基因位点的符号。这样B6·AKR也可以写作B6-H-2k。对于最近培育的同类系,小鼠遗传标准命名委员会建议同时采用两种方式,这样B6·AR或B6-H-2k应该写为B6·AKR-H-2k。
如果进一步研究嵌合体小鼠的其它性状,还可以发现嵌合体小鼠将双亲的各种不同的性状都嵌合起来了。例如Mintz(1967,1971)证明用DBA/2和C3H两种不同品种的近交系所嵌合的后代具有不同的异柠檬酸脱氢酶(IDH-1)或苹果酸脱氢酶(MDH),利用电泳技术可以将它们彼此分开。除了双亲具有的酶以外,嵌合体内有时还可以发现第三种酶,即双亲的“杂种”或异多聚酶。以上的工作对于细胞水平的遗传性研究是具有重要意义的,受到人们越来越多的重视。
嵌合体小鼠近年来已应用于细胞和组织的动力学研究,如研究小肠上皮细胞移行规律及其定位等。
2.单亲纯合双倍体动物的育成。单亲纯合双倍动物育成技术又称为雌核发育技术。这是一种相当于植物中由花粉培育成纯合双倍体植株的技术,其结果都是培育出具有两套完全一致的染色体及基因的个体。
众所周知,不论在生产上还是在科学试验中都需要纯系动物。为了得到一个纯系动物,一般要花费数年时间,甚至一生的光阴。既使对世代周期较短的小鼠,采用了最有效的兄妹交配方法,还需要连续兄妹交配20代以上才能获得纯系动物,这也要几年时间。如果采用单亲纯合双倍体育成的技术来育成一个纯系动物,则将大大地缩短育种周期,小鼠只需三周,牛也只需几个月。具体方法见图3-6。

图3-6 单亲双位体动物育成(雌核发育)
首先将两个不同品系的的近交系进行杂交(例如SJLXBL/10),交配后,在精核与卵核尚未融合之前,从母鼠子宫内冲取受精卵并用极细的吸管将雄核去掉。然后在细胞松驰素B的处理下使雌核加倍,形成二倍体细胞,二倍体细胞在体外继续培养到胚囊期后,移植到养母的子宫内使胚胎继续发育,直至出生。一般选择毛色与双亲不同的品系作为养母,以易于区分。本例中是选择毛色为野生色的BL/10XSJL杂交鼠作为养母的。养母本身没有经过交配,因而出生的小鼠均为移植进去的胚胎发育而成,它表现出亲母鼠纯合的性状。若出生的新生鼠表现为野生色,则说明实验技术失败,并未剔除精核。
单亲纯合双倍体技术也已在鱼类和家兔中获得成功。
3.细胞核移植系的产生。在单亲纯合双倍体育成技术中已提到了核移植技术,但这仅是该技术的初步。若能将受精卵中的精核,与卵核都移出,然后再将来自其它细胞新核移入受精卵,并能顺利地完成新胚胎的正常发育,核移植技术就算成功了。以前,完整的核移植技术仅在两栖类和昆虫中取得成功。1981年日内瓦大学的伊尔曼斯(K.Illmensee)和美国杰克逊研究所的霍普(P.C.Hoppe)首次在小鼠中完成了核移植技术,开创了哺乳动物核移植成功的先例。具体方法见图3-7。

图3-7细胞移植系的产生
(1)将内细胞块细胞里的核用吸管吸出。
(2)将取出的核输到另一个精核与卵核尚未融合的受精卵内。
(3)吸出该受精卵本身的精核与卵核。
(4)受精卵在体外发育到胚胎细胞。
(5)将胚盘细胞移植到白色的养母小鼠的子宫内。
(6)出生的小鼠长出灰毛,说明细胞移植系实验成功。
(1)首先用灰色的小鼠品系进行雌雄交配(也可用其它毛色的品系),从母体的子宫内获得胚盘细胞。胚盘细胞包括营养外胚叶细胞和内细胞块细胞两种,后者将来可形成胚胎。若错误地取了营养外胚叶细胞就要导致实验失败。
(2)用极细的吸管取出内细胞块细胞的细胞核,并将它注到刚受精的、但卵核与精核尚未融合的黑色小鼠的受精卵内。
(3)从黑色小鼠的受精卵内取出吸管时,将黑色鼠受精卵原有的卵核和精核吸出。
(4)将处理过的受精卵在体外培养到胚盘细胞期,再将它移植到白色的养母小鼠的子宫内,让它继续发育。
(5)产出的新生小鼠必须会长出灰色的毛,这就是细胞核移植鼠。
由于主要的遗传物质存在于细胞核中,所在核移植的受体将具有与核移植供体完全相同的遗传基础。因此,细胞核移植系技术在动物育种工作中具有十分重要的意义。因为它象植物中的组织培养那样,可以利用一头优质的动物(例如高产的奶牛)复制出与它生得完全一样的成千上万的细胞核移植系动物。
(二)实验小鼠染色体工程进展
所谓染色体工程是以染色体为单位进行有意识的切割、修补或成条、成套地增加,将人类需要的遗传性状集中在一起,创造出新的物种。以往的染色体工程一般均采用物理的或化学的方法。在高等动物中,由于多倍体或者染色体严重缺失与重复的个体都不容易成活,所以高等哺乳动物的染色体工程目前均是在细胞水平上采用细胞融合技术进行。
Harris和Watkins(1965)首先制备了包含小鼠和人类细胞的种间异核本。他们将由宫颈癌组织培养出来的人类HaLa细胞和小鼠细胞(生长在腹膜腔中Ehrlich小鼠肿瘤细胞)置于同一培养液中,加入经紫外线灭活了的仙台病毒(Sendai Virus,是粘病毒的副流感群中的一种)。在仙台病毒的作用下使两种细胞发生融合。融合细胞开始有两个来自不同物种的细胞核,称为异核体,这一点可以通过同位素标记的方法加以证实,事先他们用氚标记的胸苷来标记HaLa细胞的细胞核,而在一个异核体细胞内,可以发现标记的和不标记的两个细胞核。异核体的两个核融合后就可以继续进行细胞分裂,并形成单核细胞系。杂种细胞大,容易认别。它在以后的不断的分裂过程,人类的一套染色体和小鼠的一套染色体并不是都随着细胞的分裂进行复制的。在正常的条件下,人类的染色体将随着细胞的分裂而逐渐丢失。这种丢失是随机的。所以最后形成的杂种细胞往往都是带有一整套小鼠染色体加上不同数目和不同编号的人类染色体(或带有丝粘的片断),由于现代染色技术发展,使人们不仅可根据染色的结果很容易地区分人类染色体与小鼠染色体,而且可以确定在杂种细胞中留下来的是哪一条或者哪几条人类染色体。
如果使用的小鼠细胞是营养缺陷型细胞,即细胞本身不能合成某种必需营养物,只有在培养液中添加了这种营养物质后细胞才能正常地生长。小鼠营养缺陷型根据其所需的不同种营养物有许多不同的种类。如果将某种小鼠营养缺陷型细胞与人类正常细胞融合,形成杂种细胞。这时小鼠的营养缺陷可由于人类染色体上的有关基因的存在,该种生存必需营养物得到弥补,杂种细胞在不添加该营养液中的也能正常生长。经过一段时期培养,保留下来的细胞除带有一整套小鼠染色体外,至少还稳定地保留了一条与该营养物合成有关的人类染色体。如果我们使用多种营养缺陷型细胞作为遗传标志,就可有得到许多不同的杂种细胞,通过分析比较,就可能确定在人类的遗传中那种营养物质合成的基因在哪一条染色体。我们不仅可以用营养缺陷基因作为标志,还可以采用生化、免疫学和医学有关的基因作为标志基因,这样就可以把决定嘌呤酶的基因,决定各种碳水化合物、氨基酸以及脂肪代谢的基因,氨基酰-tRNA合成的基因,遗传性疾病的基因(例如白内障、指甲髌骨发育不全),对白喉和脊髓灰质炎等毒素易感性基因,细胞表面抗原基因和多肽酶基因等作为标志基因,结果就可以完成对更多的基因染色体定位工作。现在已定位在人类染色体的基因数目已超过了210个,对人类遗传学的研究作出了很大的贡献。
(三)实验小鼠基因工程进展
超级小鼠的产生。1982年末Palmiter和Brister报导,他们把小鼠MT-1基因的启动子与大鼠生长激素(GH)基因结合,制成融合基因(MGH),然后将MGH插入到大肠杆菌的质粒中,扩增,备用;把收获的MGH基因注入小鼠受精卵的雄性前核;然后将这些受精卵移植到养母小鼠的子宫内,直至分娩。
结果在出生的21只小鼠中有7只带有MGH基因,6只小鼠的每个细胞均有两个以上的MGH(多的可达35个)。在携带MGH的小鼠肝细胞中检出的MGH-mRNA数目高达3000个血液中生长激素浓度明显增高,表明MGH基因已得到表达。含有两个以上MGH基因的6只小鼠,生长迅速,个别的为正常小鼠的两倍,成为所谓的“超级小鼠”。含有MGH基因的小鼠与普遍小鼠杂交的结果,子代19只小鼠中有10只含有MGH基因。MGH基因可以从亲代向子代递这一事实说明MGH基因已整合到小鼠染色体上去了。
最近也有人报导了利用人类的生长激素基因使小鼠成为超级小鼠的。超级小鼠的育成,开创了基因工程在哺乳类动物中获得成功的首例。它对于医学和畜牧业的发展有着极其深远的意义和广阔的前景。
第六节 常用近交系动物品系特征
一、常用近交系小鼠品系特征
这里着重介绍目前国际上公认的主要品系和我国常用的品系。
(一)A系
1.形成
(1)起源:1921年Strong从Cold Spring Harbor albino和Bagg albino杂交的后代中近交培育而成。
(2)亚系:A,A/He,A/J,A/WySN等。
(3)近交代数:116代(Strong,1972)
2.毛色和H-2基因:aa、bb、cc(白色),H-2a(A,A/He,A/SnSf,A/WySN)。
3.主要特性:
⑴免疫学特征:44%6月龄母鼠。红斑狼疮细胞和抗核抗体阳性。缺乏补体C5。对抗原注射的免疫应答良好,但对8周龄小鼠腹腔注射0.1-0.05mg人丙种球蛋白或兔丙种球蛋白的离心上清液,2周后再皮下注射0.25mg,便出现免疫耐受性。巨噬细胞的碳粒清除率高。能及时排除抗原体复合物,不易患免疫复合物病。干扰素产量低。
⑵形态学特性:初生仔鼠的7.6%有唇裂,0.5%有后肢多趾症。
⑶生理学特性:血压低,肾上腺贮存脂质较多。245日龄以前易引起中度听觉性痉挛发作,对X线照射高度敏感。老龄鼠可自发淀粉变症。
⑷癌发生率:肺癌母鼠5-6%(A,A/He)、30%(A/J)。肝癌10.8%(A/He)。乳腺癌繁殖母鼠为80%(A)、40%(A/He)、25%(A/J)。肺组织对化学致癌物甲基胆蒽敏感。
⑸老化及寿命:老龄鼠多发肾脏病,有时出现心脏病。寿命平均400天。SPF动物雄雌分别为512天和588天。
(二)AKR系
1.形成:
⑴起源:最早是洛克菲勒大学以随机交配维持的动物,1928-1936年Furth作为白血病易发群维持着。Rhoades将其兄妹交配了9代,之后Lynch进行到21代。
⑵亚系:AKR,AKR/A,AKR/J,AKR/N等。
⑶近交代数:128代(美国N.I.H1973)。
2.毛色和H-2基因:aa,BB,cc(白色),H-2k。
3.主要特性:
⑴免疫学特征:缺乏补体C5。容易诱发免疫耐受性。用聚乙稀吡咯烷酮一次免疫8-10周龄小鼠产生抗体能力低,可能因B细胞分化能力低。但二次免疫后抗体产生良好。对白血病因子敏感。对百日咳组织胺易感因子敏感。干扰素产量高。
⑵生理学特性:血液过氧化氢酶活性高,肾上腺质贮存少,类固醇浓度低。白细胞增多症从5~6个月后的小鼠即可发现,但经常在8~9个月以后有80~90%有(雌鼠90%,雄鼠65%)白细胞增多症,但很少见到淋巴细胞而经常见到髓性白细胞,最常见到者为白细胞母细胞的增多。
⑶癌发生率:淋巴性白血病为60-100%。
⑷寿命:SPF动物雄雌分别为35天和312天。
⑸饲养注意点:在普通环境中繁殖率低。在无菌和SPF环境中繁殖良好。生育期短。应使其在2-5月龄间繁殖。
(三)BALB/c系
1.形成
⑴起源:Bagg从1913年开始维持的白化小鼠,Dowell从1923年开始近亲交配,1923年Snell得到其第26代,此后便传播各地。
⑵亚系:BALB/c,BALB/ccd,BALB/cJ,BALB/cAnN等。
⑶近交代数:157代(美国N·I·H1973)。
2.毛色和H-2基因:AA、bb、cc(白色),H-2d。
3.主要特性:
⑴免疫学特性:易患慢性肺炎。多数个体于6月龄以后出现免疫球蛋白过多症。主要是IgG1和i gA量的增加,免疫球蛋白的绝对量依饲养条件而异。腹腔注射矿物油后可引起浆细胞瘤。发生率0-60%,依亚系及饲养条件而异。较难诱发免疫耐受性。
⑵生理学特性:血压较高。网状内皮系统器官与体重之比较大。大部分雄鼠在20月龄前出现脾脏淀粉样变性。对X线照射极为敏感。对鼠伤寒沙门氏菌C′5敏感,对麻疹病毒中度敏感。
⑶癌发生率:乳腺癌发生率低(3%)。常发生肾上腺及肺有癌症。雌鼠的肺癌发生率为21%,网状细胞瘤为8%,血管瘤为6%,淋巴肉瘤为4.5%。对致瘤因子敏感。
⑷老化及寿命:老龄鼠出现心脏病、多发动脉硬化症。SPF动物雄雌的寿命分别为509天和561。
4.饲养注意点:生育期较长(至一年龄)容易饲养繁殖。最适温稍高,为26-27℃。因为体型较细,除了日常饲料外,每日早晚每只喂1cm3白面包,每星期喂二次木鱼(日本传统加工食品)。稍肥后较易繁殖。
因为易患肺炎,最好不要和其它近交系小鼠同室饲养。不使其它近交系小鼠发病的环境病原菌易使BALB/C患病。
(四)CBA系
1.形成:
⑴起源:1920年Strong从雌性Bagg albino和雄性DBA杂交后代中近交培育而成。
⑵亚系:CBA/Br,CBA/Ca,CBA/J,CBA/St,CBA/H等。
⑶近交代数:159代(杰克逊实验室1973)。
2.毛色和H-2基因:AA、BB、CC(野生色),H-2k。
3.主要特征:
⑴免疫学特性:易诱发免疫耐受性。
⑵形态学特性:18%的动物出现下颚第三臼齿缺失。
⑶生理学特性:血压较高。对维生素K不足高度敏感。连续注射酪蛋白后较C3H更易引起淀粉样变症。
⑷癌发生率:雌鼠乳腺癌60-65%,雄鼠肝癌为65%。
⑸寿命:SPF动物的雄雌鼠分别为486天和825天。
(五)C3H系
1.形成
⑴起源:与CBA同一起源。
⑵亚系:C3H/Bi,C3H/He,C3H/HeJ,C3H/St,C3HeB/FeJ,C3H/DiSn,C3H/Sf等。
⑶近交代数:138代(杰克逊实验室1973)。
2.毛色和H-2基因:AA、BB、CC(野生色),H-2k。
3.主要特性:
⑴免疫学特性:在IgG的亚类中IgG1和IgG2a为高值,IgG2b为低值。IgG1量增加到生后第三个月,以后开始下降,因为IgG2a变动不大,所以9月龄时与IgG1相比为高值。IgG1b值IgA为100-200μg/ml的低值,无变动。IgM伴随增龄有所增加。无菌饲养的IgM量为普通饲养的2倍以上。
较易诱发免疫耐受性。C3H/Hel对脂多糖非特异性活化B细胞的作用呈低反应性,其它C3H为高反应鼠。对鼠伤寒沙六氏菌补体5有抗力。
⑵生理学特性:红细胞及白细胞数较少。皮下注射5%酪蛋白0.5ml,5次/星期,3周后全部患淀份样变症。血液中氧化氢酶活性高。
⑶癌发生率:乳腺癌繁殖用雌鼠为85-100%。是通过乳汁感染而不是胎盘感染。白血病雌雄分别为0.5%和14%。肝癌雌雄分别为0%和10%。14月龄雄性小鼠肝癌自发率为85%。
⑷饲养注意点:较易饲养。繁殖用雌鼠从较早期(约5月龄)便有乳腺癌发生。患乳腺癌后幼仔哺育不良。哺仔下痢症感染率高。雄鼠对氨气,雄鼠对氨气,氯仿,松节油等气体特别敏感,且死亡率高。该鼠雌性不强。
(六)C57BL/6系
1.形成:
⑴起源:1921年Little由Abby Lathrop得到动物后开始近亲交配,育成数个近交系。以57号母鼠和52号公鼠交配为起源者标为C57,C77中毛色固定为巧克力色者称为C57BR,固定为黑色者称为C57BL。
⑵亚系:C57BL/6,C57BL/6J,C57BL/6N等。
⑶近交代数:114代(杰克逊实验室1973)
2.毛色基因:aa、BB、CC(黑色)
3.主要特性:
⑴免疫学特性:IgG在20月龄前缓慢增加,IgG2b为高值,IgG为低值。无菌饲养较普通饲养者IgG绝对量低。IgG为高值,有的个体12个月龄后可超过800μg/ml。无菌饲养的IgM较高。细胞免疫力随增龄较少降低,可能与自发肿瘤较少有关。较易诱发免疫耐受性。干扰素产量较高。对百日咳易感因子(Pertussis HSF)敏感。
⑵形态学特性:新生仔中雌性的16.8%,雄性的3%为小眼或无眼症。0.6%出现后肢多趾症。
⑶生理学特性:嗜酒精性高。肾上腺贮存脂质少。将雄鼠皮肤移植至同系雌鼠,20天后出现排拆。此为组织相容性原中存在雄性抗原,C57BL较其它系小鼠为显著。注射酪蛋白后易引起淀粉样变症。用考的松可诱发出20%腭裂,对结核杆菌敏感,对鼠痘病毒有一定抗力。
⑷癌发生率:乳腺癌少发(0-1%)。用致癌难以致癌。老龄鼠淋巴瘤自发率为20~25%;雌鼠白血病为7~16%,经照射后肝癌发生率高。
⑸寿命:最长达1200天。平均雌鼠692天,雄鼠676天。
4.饲养注意点:繁殖率稍低,给予高脂饲养可好转。离乳时的幼仔可出现原因不明的脱毛,在每次饲喂时,除了固型饲养外给予少量成鸡用混合颗粒料可防止离乳时的脱毛。
(七)DBA/1系
1.形成:
⑴起源:为1909年Little在毛色遗传实验中使用的动物。1929-1930年间培育成几个品系,现存的有DBA/1和DBA/2
⑵亚系:DBA/1,DBA/1J等。
⑶近交代数:83代(杰克逊实验室1973)
2.毛色基因:aa、bb、CC、dd(淡巧克力色)
3.主要特性:
⑴免疫学特性:对实验性结核感染的易感染性高。
⑵生理学特性:红细胞数多。繁殖用动物的大部分出现心脏钙质沉着。
⑶癌发生率:经产母鼠的乳腺癌为61.5%。白血病为8.4%。
⑷寿命:SPF动物的雄雌鼠分别为487天和684天。
(八)DBA/2系
1.形成:
⑴起源:参阅DBA/1。
⑵亚系:DBA/2,DBA/2J等。
⑶近交代数:116代(美国N.I.H1973)。
2.毛色基因:aa、bb、cc、dd(淡巧克力色)。
3.主要特性:
⑴免疫学特性:在普通饲养条件下三月龄鼠血清免疫球蛋白量为1000μg/ml左右,仅相当C57BL/6,C3H/He和BALB/C的1/2。其中,IgM值较高,为221.6μg/ml。而IgG为低值,仅相当于其它品系小鼠的1/2-1/4。在IgG的各亚类中,IgG1高值,IgG2b最低。缺乏补体C5。DBA/2和CBA的杂交一代于18月龄时发生浆细胞性白血病,在血清中测出与BALB/C浆细胞瘤相同的M成分。
⑵生理学特性:红细胞多。血压较低。听源性癫痫发作率在35日龄时为100%,55日龄时为5%。维生素K缺乏,氯仿和氧化乙烯引起的死亡率高。肾上腺脂质贮存少,心脏有钙盐沉着。嗜酒性低。对百日咳组织胺易感因子敏感,对疟原虫感染有一定抗力,对鼠斑疹伤寒补体5有抗力。
⑶癌发生率:经产母鼠乳腺癌发生率50-60%,雄雌鼠白血病分别为8%和6%。雌雄鼠中均有淋巴瘤生长。
⑷寿命:SPF动物的雌鼠为719天,雄鼠为629天。
⑸饲养注意点:产仔数少,不易哺育,所以较难繁殖。
(九)NZB系
1.形成
⑴起源:1930年Hill从Otago医科大学引起动物,1948年开始近亲交配。同一起源的还有NZO、NZW、NZX和NZY。
⑵亚系:NZB,NZBfC3H/HeLac。
⑶近交代数:96代(Bielschowsky 1973)。
2.毛色基因:aa,BB,CC(黑色)
3.主要特性:
⑴免疫学特性:
a、自身免疫性溶血性贫血症。在4-5月龄以前Ht值平均为44%,以后伴随增龄而下降,14月龄为32%,雌鼠在6月龄后,雄鼠在8月龄后出现Coombs氏试验阳性。伴随增龄阳性比率增高,10月龄为50%,12月龄以后达100%。从6月龄起出现抗核抗体。老龄小鼠出现红斑狼疮细胞。
b、血清免疫球蛋白:血清免疫球蛋白量异常之高,特别是IgM和IgG量递增显著。IgM量在一生中递增,其递增与性别、病况及Coombs氏试验无关,并不因无菌饲?
■[此处缺少一些内容]■
4.8%,其后报导了胸腺病,淋巴性白血病,浆细胞白血病及恶性淋巴瘤等。淋巴性白血病的发生率为7%。常有未成熟期发病者,一般多在3-8月龄发病。
⑵寿命:SPF的雌鼠为441天,雄鼠为459天。
4.饲养注意点:哺育不易,繁殖极为困难。有肥胖的倾向。饲养繁殖的主要困难是母鼠的受孕期仅在4月龄之前,较其它近交系为短。并且在产仔后,有咬死自产仔鼠的习性。
(十)SJL/N
1.形成:
⑴起源:由Swiss-Webster小鼠发展面来,1938~1943年间到Jackson研究室。1955年开始近交繁殖,1969年到美国NIH,按近交繁殖保种。
⑵近交代数:F14(1973年)。
2.遗体组成:C/C:H-2s,Tlaa·lg-1b,Hbac·HbbS。
3.主要特性:
⑴一年以上的小鼠中有类何杰金氏病的多形细胞性网织细胞肉瘤。
⑵对全身性X线照射抗力强。
⑶对百日咳组织胺易感因子敏感。
⑷为Wanebo网织细胞肉瘤的宿六。
(十一)AFB系
1.形成:该品系未列入国际名单,是苏联培育的,在苏联资料中记载。但未介绍是如何培育的,可能与AKR有关,我国血研究在1958年从苏联引起,在1971年由天津引入医科院分院,现已40多代。
2.主要特性:该品体系质弱,体形中等,繁殖力弱,二月龄体重20~22克,很少有自发性肿瘤。
该品系为白色细胞增多症的白色小鼠。从5~6个月后的小鼠即可出现白细胞增多,但经常在8~9个月,以后有90%的雌鼠和65%的雄鼠可以发生白细胞增多症,但很少见到淋巴细胞而是经常见到髓性白细胞,最常见到者为白细胞母细胞的增多。
(十二)C3HA系
1.形成:Pogosianj于1943年用C3H和A杂交而育成。因此该鼠为野灰色但尾尖部总带有白色,有时白色多些,有时少些,但无论如何消除不掉。我国在1958年时血研所从苏联引进,当时称“苏乙”品种,1971年由血研所引入中国医科院分院,该鼠体形小,二月龄时体重为17~20克,但繁殖力强。
2.主要特性:
⑴乳腺癌发病率高,生育过或未生育过的雌鼠的癌肿发病率均为90%左右。
⑵经常可以见到肺腺癌与性别无关。
⑶我国饲育时有30%的自发性乳腺癌,并对致肝癌物质敏感。
(十三)我国培育的近交品系
见表3-5。
表3-5 我国培育的小鼠近交品系
| 品系名称 |
培育单位及起源历史 |
毛色(基因型) |
主 要 特 性 |
| 中国1号(C-1) |
1955年中国医学科学院实验医学研究所,将昆明小鼠经近亲交配20代以上而育成。 |
白 化 |
繁殖力中等,二月龄体重17克,肿瘤自发率低。 |
| 津白1号(TAⅠ) |
1955年天津医学院昆明市售白化小鼠经的近亲交配而成。 |
白化(aabbcc) |
繁殖力中等,二月龄体重20-25克,肿瘤自发率低。 |
| 津白2号(TAⅡ) |
1963年天津医学院将昆明种小鼠经近亲交配20代以上而育成。 |
白化(aabbcc) |
高乳腺癌,繁殖力中等。 |
| 615 |
1961年中国医学科学院输血和血液研究所将普通小白鼠与C57BL/血研究杂交所生子代经亲代交配20代以上而育成。 |
深褐色(aabbcc) |
肿瘤发生率为10-20%(♀♀乳腺癌,♂肺癌)对津638白血病毒敏感 |
| AMMS/1号 |
1974年军事医学院科学院将昆明种小白鼠经亲代交配20代以上而育成。 |
白化(aabbcc) |
对炭疽弱毒株比较敏感,地骨髓多向性造血干细胞测定较规律。 |
二、常近交系大鼠品系特征
(一)ACI系
1.形成:
⑴起源:1926年由哥仑比亚大学肿瘤研究所Curtis和Dunning培育。1945年Heston繁殖30代,1950年美国NIH繁殖41代,最后由Dunning(或NIH)育成。
⑵近交代数:F>100。
2.毛色:黑色,但腹部和脚白色(a,hi)。
3.主要特性:
⑴生理、生化学特性:血清甲状腺素含量低。收缩压低。雌鼠苯胺的肝脏代谢率高。戊基巴比妥纳的LD50值(为120mg/kg)高。
⑵形态学特性:28%雄鼠和20%雌鼠患有遗传缺陷病,有时一侧肾发育不全或肾囊肿,与子宫角缺陷或同侧睾丸萎缩有关。这些畸形,都与一种多基因遗传方式有关。
⑶癌发生率:雄鼠自发性肿瘤:睾丸为46%,肾上腺为16%,脑垂体为5%,皮肤和耳道以及其他类型为6%。雄鼠自发性肿瘤:脑垂体为21%,子宫为13%,乳腺为11%,肾上腺和其他类型为6%。34~37月龄老年雄性小鼠,自发性前列腺癌17%。该品系小鼠可使M-C961、970、R3234肿瘤移植生长,吸收中等剂量的乙烯雌酚后乳腺肿瘤增加。
⑷饲养繁殖:繁殖力差,仔鼠矮小,胚胎死亡率高,取决于雄鼠基因型。早期胚胎死亡率11%,先天性畸形发生率10%。
⑸寿命:需长时间驯养,对新环境的适应时间也长。雄鼠平均活存时间113周,雌鼠108周。
(二)AGUS系
1.形成
⑴起源:Gustafsson用动物株(Sprague-Dawley)培育的无菌动物,1948年繁殖10代,到1968年实验动物中心Garshalton繁殖26代。
⑵近交代数:F35。
2.毛色:白化(a.c.H)。
3.主要特性:易感染实验过敏性脑脊髓炎,对溶组织阿米巴的感染有相对抗力。尽管动物易受环境影响,但具有良好的繁殖能力。约有1~4%无尾年幼大鼠无繁殖力。
(三)AS系
1.形成:
⑴起源:1930年Otago大学用从英国引入的Wistar大鼠繁殖,它可能是GH品系,因而具有组织相溶性。
⑵近交代数:F60+。
2.毛色:白化(C)。
3.主要特性:血压不如GH品系那样高,但仍较正常血压要高。易感染实验过敏性脑脊髓炎,对自家免疫性肾小球肾炎敏感,这与其组织相溶性有关。具有良好的繁殖力。
(四)BN系
1.形成:
⑴起源:1958年由Silvers和Billingham用D.H.King和P.Aptekman培育的棕色突变型动物近交繁殖而来。
⑵近交代数:F35。
2.毛色:棕色(a、b、h1)。
3.主要特性:
⑴免疫学特性:对实验过敏性脑脊髓炎有抗力。对自家免疫性复合性肾小球肾炎也具抗力。⑵生理学特征:平均31月龄大鼠心内膜疾病的发生率为7%,先天性高血压为30%,对戊基巴比妥钠中度敏感,其LD50为90mg/kg。
⑶癌发生率:雌雄大鼠上皮瘤的发生率分别为28%和2%。雌鼠输尿管肿瘤20%,雄鼠6%。雄鼠膀胱癌自发率35%,胰腺腺瘤15%,脑垂体腺瘤14%,淋巴网状组织肉瘤14%,肾上腺皮质腺瘤12%,髓质性甲状腺瘤9%,肾上腺嗜铬细胞瘤8%。雌鼠脑垂体腺瘤26%,输尿管癌22%,肾上腺皮质腺瘤19%,子宫颈肉瘤15%,乳腺纤维腺瘤11%,胰腺腺瘤11%。
⑷寿命:该品系繁殖力低。雄鼠平均寿命29个月,雌鼠为31个月。
(五)BUF系
1.形成:
⑴起源:系美国布法罗H·Morris株,1946年由Heston培育,1956后NIH繁殖了10代。
⑵近交代数:F58。
2.毛色:白化(C)。
3.主要特性:
⑴免疫学特性:36周龄的雄鼠有54%发生自家免疫性甲状腺炎,大于一年龄的动物有26%发生自发性自家免疫性甲状腺炎和甲状腺单核细胞浸润。食入3-甲基五环碳氢化合物能自发地发生自家免疫性甲状腺炎,而新生期胸腺切除后其发生率几乎达到100%。对绵羊红细胞缺乏免疫反应。
⑵生理学特性:龋齿发生率低。乙基吗啡的肝脏代谢率低,而苯胺的肝脏代谢率高。产前早期死亡率18%,晚期死亡率11%。繁殖力和仔鼠大小中等。未发现先天性畸形。
⑶癌发生率:Yoshida氏腹水瘤16%,有Morris氏肝癌和脑垂体肿瘤生长。在老年动物中,早期自发性脑垂体瘤30%,肾上腺皮质瘤25%,患有甲状腺癌的25%,两年龄大鼠活存率为58%。
(六)COP系
1.形成:
⑴起源:1921年由哥仑比亚肿瘤研究所Curtis培育。
⑵近交代数:F73。
2.毛色:头部被毛呈黑色头巾状(a,h)。
3.主要特性:所产生自发性胸腺肿瘤。吸收小剂量乙烯雌酚能使动物产生膀胱结石和乳头瘤而死亡。对乳腺瘤有抗力,脑垂体小。对囊尾蚴虫有抗力。允许IRS4337和前列腺癌R3327肿瘤移植生长,而后者是人类前列腺癌的动物模型。平均寿命为20±0.2月。
(七)F344系
1.形成:
⑴起源:1920年由哥仓比亚大学肿瘤研究所Curtis培育,1949年引入Heston之后又到NIH,1950年Bethesde将其繁殖51代,最后完成繁殖的不是Dunning就是NIH。
⑵近交代数:F86。
2.毛色:白化(a,c,h)。
3.主要特性:
⑴免疫不特性:原发和继发性的脾脏红细胞的免疫反应性低。特殊活动性能高,但与远交系Sprague-Dewley大鼠比较,其NADPH-细胞色素C还原酶的诱发力低。
⑵生理学特性:旋转运动性低。雌雄鼠肝结节状增生的发生率为5%。血清胰岛素含量低。雄鼠乙基吗啡和苯胺的肝代谢率高。对高血压蛋白盐的产生有抗力。脑垂体大。对囊尾蚴虫敏感,乙烯雌酚吸收快并易引起死亡,尸检时脂肪肝很常见。戊巴比妥钠的LD50低,为70mg/kg。肾脏疾患发生率低。
⑶癌发生率:雌鼠乳腺癌自发率为41%,脑垂体腺腺瘤为36%,雌鼠乳腺癌为23%,脑垂体腺瘤为24%,睾丸间质细胞瘤为85%。甲状腺癌22%,单核细胞白血病24%。雌鼠乳腺纤维腺瘤9%,多发性子宫内膜肿瘤12%。在无菌条件下肿瘤的发生率:雄鼠白血病26%,而雌鼠为36%,雄鼠乳腺瘤12%,而雌鼠为20%,雄鼠可见其他肿瘤9%,而雌鼠可见5%。该品系大鼠可允许下列肿瘤移植生长:Dunning氏肝癌、肝癌LC-18、Novidoff肝癌、乳腺癌HMC和R-3230、脑垂体瘤MtT和MtTf4、Walker256类癌肉瘤,Dunning白血病,白血病HLF、IRC-741和R3149,淋巴肉瘤R-3251,乳腺纤维瘤F-609,纤维肉瘤R-3244,肉瘤IRS9802和R13259,子宫肉瘤F-529和白血病R-3323、3330、3399和3432。
⑷寿命:雄鼠平均31月,雌鼠水均29月。
(八)GH系
1.形成:
⑴起源:1930年来源于英国报导的Wistar大鼠,由牛津大学医学研究所增育。1955年Snirk开始研究选择高血压大鼠,繁殖了许多品系,该品系是其中的一种而且在分类上与AS品系有关联。
⑵近交代数:F31。
2.毛色:白化(C)。
3.主要特性:高血压,有心肌肥大和血管疾病,与正常血压的品系的相比,心率快20%,体脂肪含量较低,而心脏重量大50%。GH品系(但不是SHR)遗传性高血压可能与肾前列腺素的分解代谢有关。与SHR品系有关的特征已由Simpson等人作了报导。
(九)IS系
1.形成:
⑴起源:来源于野生雄鼠和Wistar雌鼠,自1968年以来,人们就开始用同种亲缘动物交配方法培育该品系动物。
⑵近交代数:F23。
2.毛色:刺鼠(+)。
3.主要特性:多见有胸腰椎骨畸形并导致脊柱后侧凸,使脊髓管阻塞,脊髓受压,畸形好发在第12胸椎到第6腰椎的部位,通常有两上到四个脊椎受累,但有时可多到7个,而且某种个体的畸形发生率高达90%。
(十)LEW系
1.形成:
⑴起源:来源于Wistar原种,最先由Lewis繁殖,1954年Aptekman和Bogden繁殖到20代,到1958年Silvers繁殖到31代。
⑵近交代数:F67。
2.毛色:白化(a,h,c)。
3.主要特性:
⑴免疫学特性:接种豚鼠髓磷脂硷蛋白后,对实验过敏性脑脊髓炎敏感。极易感染诱发自家免疫性心肌炎。对诱发自家免疫性复合性肾小球肾炎敏感(这与主要组织相溶性复合物有关)。易感染实验过敏性脑炎和药物诱发的关节炎。
⑵生理学特性:血清甲状腺素高,血清胰岛素和血清生长激素高。动物的肥胖取决于饮食的高脂肪物的含量。雌鼠乙基吗啡的肝脏代谢率高。
⑶癌发生率:常见淋巴瘤,肾肉瘤,纤维肉瘤MC-39,ML-1,ML-7,Lewis10瘤和Lewis3肉瘤。
⑷饲养繁殖:易驯养,繁殖率高。二年龄大鼠的存活率为26%。
(十一)LOU/c LOU/M系
1.形成:
⑴起源:可能来自Louvain大学Wistar株,由Bazin和Backers培育。28种类似的LOU/C品系动物可用作高发性浆细胞瘤的研究。两者的组织相溶性相同,这就是LOU/M雄鼠接受LOU/C动物的皮肤移植的基础。
⑵近交代数:F20+?。
2.毛色:白化(?)。
3.主要特性:LOU/C和LOU/M特性参看第十章第二节。
(十二)M520系
1.形成:
⑴起源:1920年由哥仑比亚大学肿瘤研究所Crutis培育,1949年Heston繁殖49代,1950年NIH繁殖到51代。
⑵近交代数:F86。
2.毛色:白化(a,c,h)。
3.主要特性。
⑴生理学特性:收缩压低。苯胺的肝脏代谢率低(雄鼠90%,雌鼠70%),但乙基吗啡的代谢率高。繁殖力强,仔鼠中等大小。易感染囊尾蚴病。极易感染肾炎。
⑵癌发生率:NIH品系肾上腺髓质肿瘤发生率为21~25%。对2-乙酰氟胺(2-Acetylaminofluorine)诱发肿瘤敏感。胰外分泌腺瘤的发生率低。小于18月龄的动物子宫、脑垂体前叶、肾上腺髓质和皮质以及间质细胞瘤发生率在10%以内。但大于18月龄的12~50%雌鼠有子宫瘤,35%未交配雄鼠有间质的细胞瘤,65~85%动物表现出肾上腺髓质瘤、20~45%为皮质瘤,20~40%有脑垂体前叶瘤。能使Jensen肉瘤和Yoshida腹水瘤生长,75%的动物能生长7974型肝癌和130型肝癌。常见的有BICR/MI乳腺瘤、344成骨肉瘤、343癌、Harderian2226腺癌、338癌和E-2730肉瘤生长。
(十三)PETH系
1.形成:
⑴起源:1938年由Bourne引入Sidman,1946年NIH繁殖9代,以后又繁殖到18代。该品系可能与RCS品系有关。
⑵近交代数:F39。
2.毛色:粉红色眼睛,头部被毛呈头巾状(a,h,p)。
3.主要特性:携有视网膜营养不良(Retinal Dystophy)基因(re),常引起3月龄大鼠白内障。可诱发常见的晶状体反射和持续性玻璃状体膜动脉的继发反应。收缩压低。
(十四)PVG系
1.形成:
⑴起源:英国皇家科学院培育后引入Lister和Virol研究所,1946年被Glaxo引入并交繁殖。
⑵近交代数:F70+?。
2.毛色:头部被毛呈黑色头巾状(a,h)。
3.主要特性:对诱发免疫性甲状腺炎有抗力。易感染溶组织阿米巴病。Howard已经培养出染色体分带与CBA-T6品系相类似的品系。
(十五)SHR系
1.形成:
⑴起源:源于东京远交系Wistar大鼠,1963年Okamoto培育。用群体动物中患有自发性高血压的雄鼠与一只血压升高的雌鼠交配繁殖,之后进行兄妹连续交配,以获得自发性高血压动物模型。该品系中的动物有发生脑血管损伤和中风的趋势。
⑵近交代数:F23+。
2.毛色:白化(C)。
3.主要特性:高血压发生率高,而且无明显原发性肾脏或肾上腺损伤。高血压很严重,血压常高于200mmHg。心血管疾病发生率高。基因分析表明这一情况受3~4个基因控制,其中一个可能是主要的。有人认为下丘脑血压的维持是精神错乱的结果。尿嘌呤糖尿病能进一步使血压增高,但动物对抗高血压药有反应。Roba肯定该品系是筛选抗高血压反应药理最适宜的动物模型。在年幼SHR大鼠中,血浆去甲肾上腺素和多巴胺b-氢氧酶水平增加,并超过WKA品系对照大鼠。但总儿茶酚胺无明显不同,肾上腺儿茶酚胺含量减少。循环血中的促肾上腺激素水平明显高于对照品系。131I代谢率减少和甲状腺重量增加与Wistar对照鼠有关。
(十六)WAG系
1.形成:
⑴起源:源于Wistar株,1924年A·L·Bacharach培育。
⑵近交代数:F101。
2.毛色:白化(a、c、h;A、C、h或A、C、H)。
3.主要特性:
⑴生理学特性:旋转运动率高。脑GABA含低。平均31月龄大鼠心内膜疾病的发生率为4%。对铁缺乏敏感。
⑵免疫学特性:对实验过敏性脑脊髓炎有抗力。有些动物可能携带防御右旋糖酐过敏反应的隐性基因dx,对诱发自家免疫性甲状腺炎敏感。多肽类继发性抗体反应性低。
⑶肿瘤发生率:甲状腺髓质瘤27%,嗜铬细胞瘤2%,胰岛细胞腺瘤1%。31月老龄雌鼠腺瘤的发生率为69%,甲状腺髓质瘤40%,肾上腺皮质腺瘤29%,乳腺纤维腺瘤21%。
⑷平均寿命:雌鼠大于31月,雄鼠为22月。
(十七)WF系
1.形成:
⑴起源:1945年J·Furth用远交系Wistar株动物培育而成。
⑵近交代数:F27+。
2.毛色:白化(C)。
3.主要特性:
⑴生理学特性:血清生长激素含量低。对肾上腺反馈性高血压有抗力。该品系携带特殊浓度Y染色体,这种染色体可以用作细胞标记。
⑵肿瘤发生率:结肠癌雄鼠发生率为38%,雌鼠27%。雌鼠脑垂体肿瘤27%,乳腺癌21%,肾上腺肿瘤3%,白血病9%,恶性淋巴瘤7%,脂肪瘤3%。给年幼鼠食入3-甲基五环碳氢类化合物,其白血病的发生率增高,但经X射线作用后白血病则减少。
⑶平均寿命:雌鼠31月,雄鼠23月。
中国医学科学院医学实验动物研究所于1983~1988年从美国NIH、JAX,英国LAC,日本JCA等单位引入小鼠26个品系,大鼠11个品系:
小鼠品系:A/J、AKR/J-nustr、BALB/cAnN、BALB/c-nu/Jcl、BALB/c-nu/Ola、BALB/c/Ola、B6C3Fe-a/a-Dh、CBA/J/ola、CBA/N(CBA/HN)、C·B-17/IcrJ-Sc1d。C3H/He/Ola、C57BL/KsJ-dbm、C57BL/6J-ob、C57BL/6J/Ola、C57BL/6N、C57BL/6N-bg、DBA/2/Ola、DBA2N、ICR、KK、N:NIH(S)-nu、N:NIH(S)-Ⅱ-nu(N:NIH(S)-nu-Xid)、N:NIH(S)- Ⅲ-nu、NOD/Lt、SM/J、SWR/J。
大鼠品系:F344/N、F344/Ola、LOU/C-IgK-Ib、LOU-CN、LOU/MN、Nude、SD、SHR/Ola、Wistar、WKY/Ola、WN/N。

 中医世家
中医世家 浦 标 网
浦 标 网 河南大学精品课程
河南大学精品课程 图书资料室
图书资料室